Professor of Archaeology, University of Peradeniya, and Cornell Visiting Professor, Swarthmore College, Pennsylvania, Sudharshan Seneviratne delivered a speech on ‘Humanism for Peace: A Buddhist Perspective’ at the Inter Faith Dialogue on International Peace, Harmony and Co-existence celebrating Vesak at the UN General Assembly on May 16.
Diamond J. Guns, germs, and steel: the fates of human societies. W. W. Norton & Company; 2005. 83–292.
Conner SE, Kvavadza EV. Modelling late quaternary changes in plant distribution, vegetation and using pollen data from Georgia, Caucasus. J Biogeogr 2009;36: 529–45.
Byrd BF. Reassessing the emergence of village life in the Near East. J Arch Res 2005;13:231–90.
Metcalf J, et al. Synergistic roles of climate warming and human occupation in Patagonian megafaunal extinctions during the Last Deglaciation. Sci Adv 2016;23-35.
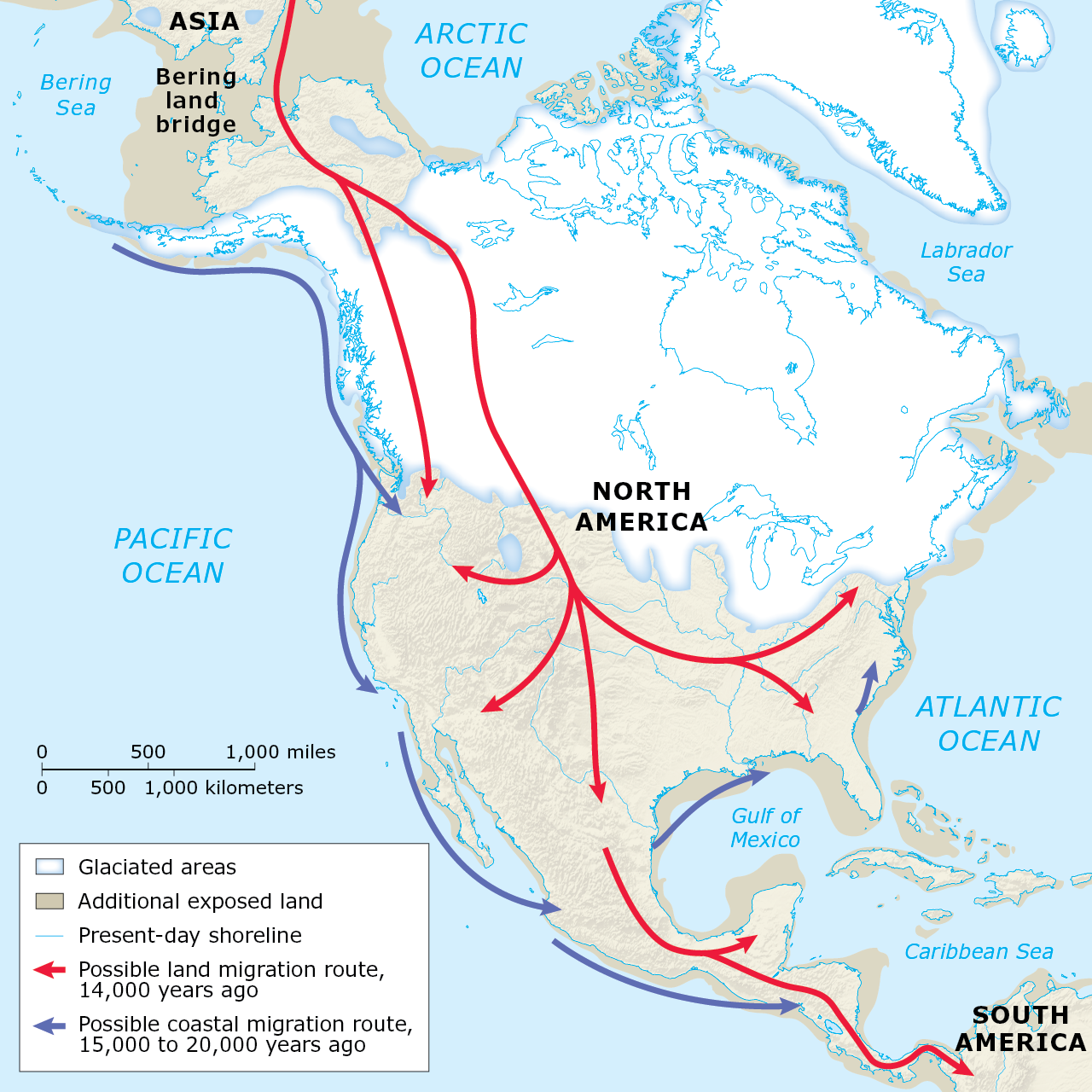
Ardelean, C. F., Becerra-Valdivia, L., Pedersen, M. W., Schwenninger, J.-L., Oviatt, C. G., Macías-Quintero, J. I., Arroyo-Cabrales, J., Sikora, M., Ocampo-Díaz, Y. Z. E., Rubio-Cisneros, I. I., Watling, J. G., de Medeiros, V. B., de Oliveira, P. E., Barba-Pingarón, L., Ortiz-Butrón, A., Blancas-Vázquez, J., Rivera-González, I., Solís-Rosales, C., Rodríguez-Ceja, M., … Willerslev, E. (2020). Evidence of human occupation in Mexico around the Last Glacial Maximum. Nature, 584(7819), 87–92.
Barker, G., Barton, H., Beavitt, P., Bird, M., Daly, P., Doherty, C., Gilbertson, D., Hunt, C., Krigbaum, J., Lewis, H., Manser, J., McClaren, S., Paz, V., Piper, P., Pyatt, B., Rabett, R., Reynolds, T., Rose, J., Rushworth, G., & Stephens, M. (2002). Prehistoric Foragers and Farmers in South-east Asia: Renewed Investigations at Niah Cave, Sarawak. Proceedings of the Prehistoric Society, 68.
Becerra-Valdivia, L., & Higham, T. (2020). The timing and effect of the earliest human arrivals in North America. Nature, 584(7819), 93–97.
Bird, M. I., Beaman, R. J., Condie, S. A., Cooper, A., Ulm, S., & Veth, P. (2018). Palaeogeography and voyage modeling indicates early human colonization of Australia was likely from Timor-Roti. Quaternary Science Reviews, 191, 431–439.
Bowler, J. M., Johnston, H., Olley, J. M., Prescott, J. R., Roberts, R. G., Shawcross, W., & Spooner, N. A. (2003). New ages for human occupation and climatic change at Lake Mungo, Australia. Nature, 421(6925), 837–840.
Bowler, J. M., & Magee, J. W. (2000). Redating Australia’s oldest human remains: a sceptic’s view. Journal of Human Evolution, 38(5), 719–726.
BOWLER, J. M., THORNE, A. G., & POLACH, H. A. (1972). Pleistocene Man in Australia: Age and Significance of the Mungo Skeleton. Nature, 240(5375), 48–50.
Braje, T. J., Dillehay, T. D., Erlandson, J. M., Klein, R. G., & Rick, T. C. (2017). Finding the first Americans. Science, 358(6363), 592–594.
Bulbeck, D. (2007). A parsimonious model for Homo sapiens colonization of the Indian ocean rim and Sahul. Current Anthropology, 48(2), 315–321.
Clarkson, C., Jacobs, Z., Marwick, B., Fullagar, R., Wallis, L., Smith, M., Roberts, R. G., Hayes, E., Lowe, K., Carah, X., Florin, S. A., McNeil, J., Cox, D., Arnold, L. J., Hua, Q., Huntley, J., Brand, H. E. A., Manne, T., Fairbairn, A., … Pardoe, C. (2017). Human occupation of northern Australia by 65,000 years ago. Nature, 547(7663).
Crabtree, S. A., White, D. A., Bradshaw, C. J. A., Saltré, F., Williams, A. N., Beaman, R. J., Bird, M. I., & Ulm, S. (2021). Landscape rules predict optimal superhighways for the first peopling of Sahul. Nature Human Behaviour, 23–29.
Davis, L. G., Madsen, D. B., Becerra-Valdivia, L., Higham, T., Sisson, D. A., Skinner, S. M., Stueber, D., Nyers, A. J., Keen-Zebert, A., Neudorf, C., Cheyney, M., Izuho, M., Iizuka, F., Burns, S. R., Epps, C. W., Willis, S. C., & Buvit, I. (2019). Late Upper Paleolithic occupation at Cooper’s Ferry, Idaho, USA, ~16,000 years ago. Science, 365(6456), 891–897.
Dillehay, T. D., Ramirez, C., Pino, M., Collins, M. B., Rossen, J., & Pino-Navarro, J. D. (2008). Monte Verde: Seaweed, Food, Medicine, and the Peopling of South America. Science, 320(5877), 784–786.
Gibbons, A. (2017). The first Australians arrived early. Science, 357(6348), 238–239.
Gilbert, M. T. P., Jenkins, D. L., Go therstrom, A., Naveran, N., Sanchez, J. J., Hofreiter, M., Thomsen, P. F., Binladen, J., Higham, T. F. G., Yohe, R. M., Parr, R., Cummings, L. S., & Willerslev, E. (2008). DNA from Pre-Clovis Human Coprolites in Oregon, North America. Science, 320(5877), 786–789.
Goebel, T., Waters, M. R., & O’Rourke, D. H. (2008). The Late Pleistocene Dispersal of Modern Humans in the Americas. Science, 319(5869), 1497–1502.
Grayson, D. K., & Meltzer, D. J. (2015). Revisiting Paleoindian exploitation of extinct North American mammals. Journal of Archaeological Science, 56, 117–193.
Harvati, K., Röding, C., Bosman, A. M., Karakostis, F. A., Grün, R., Stringer, C., Karkanas, P., Thompson, N. C., Koutoulidis, V., Moulopoulos, L. A., Gorgoulis, V. G., & Kouloukoussa, M. (2019). Apidima Cave fossils provide earliest evidence of Homo sapiens in Eurasia. Nature, 571(7766).
Hesp, P. A., Murray-Wallace, C. v., & Dortch, C. E. (1999). Aboriginal occupation on Rottnest Island, Western Australia, provisionally dated by Aspartic Acid Racemisation assay of land snails to greater than 50 ka. Australian Archaeology, 49(1), 7–12.
Hublin, J.-J., Ben-Ncer, A., Bailey, S. E., Freidline, S. E., Neubauer, S., Skinner, M. M., Bergmann, I., le Cabec, A., Benazzi, S., Harvati, K., & Gunz, P. (2017). New fossils from Jebel Irhoud, Morocco and the pan-African origin of Homo sapiens. Nature, 546(7657).
Lesnek, A. J., Briner, J. P., Lindqvist, C., Baichtal, J. F., & Heaton, T. H. (2018). Deglaciation of the Pacific coastal corridor directly preceded the human colonization of the Americas. Science Advances, 4(5), 1–10.
Malaspinas, A.-S., Westaway, M. C., Muller, C., Sousa, V. C., Lao, O., Alves, I., Bergström, A., Athanasiadis, G., Cheng, J. Y., Crawford, J. E., Heupink, T. H., Macholdt, E., Peischl, S., Rasmussen, S., Schiffels, S., Subramanian, S., Wright, J. L., Albrechtsen, A., Barbieri, C., … Willerslev, E. (2016). A genomic history of Aboriginal Australia. Nature, 538(7624).
Mallick, S., Li, H., Lipson, M., Mathieson, I., Gymrek, M., Racimo, F., Zhao, M., Chennagiri, N., Nordenfelt, S., Tandon, A., Skoglund, P., Lazaridis, I., Sankararaman, S., Fu, Q., Rohland, N., Renaud, G., Erlich, Y., Willems, T., Gallo, C., … Reich, D. (2016). The Simons Genome Diversity Project: 300 genomes from 142 diverse populations. Nature, 538(7624).
Mellars, P. (2006). Going East: new genetic and archaeological perspectives on the modern human colonization of Eurasia. Science, 313, 796–800.
Morlan, R. E. (2003). Current perspectives on the Pleistocene archaeology of eastern Beringia. Quaternary Research, 60(1), 123–132.
Neves, W. A., Hubbe, M., & Correal, G. (2007). Human skeletal remains from Sabana de Bogotá, Colombia: A case of Paleoamerican morphology late survival in South America? American Journal of Physical Anthropology, 133(4), 1080–1098.
Neves, W. A., Hubbe, M., & Piló, L. B. (2007). Early Holocene human skeletal remains from Sumidouro Cave, Lagoa Santa, Brazil: History of discoveries, geological and chronological context, and comparative cranial morphology. Journal of Human Evolution, 52(1), 16–30.
Pagani, L., Lawson, D. J., Jagoda, E., Mörseburg, A., Eriksson, A., Mitt, M., Clemente, F., Hudjashov, G., DeGiorgio, M., Saag, L., Wall, J. D., Cardona, A., Mägi, R., Sayres, M. A. W., Kaewert, S., Inchley, C., Scheib, C. L., Järve, M., Karmin, M., … Metspalu, M. (2016). Genomic analyses inform on migration events during the peopling of Eurasia. Nature, 538(7624), 238–242.
Pearce, R. H., & Barbetti, M. (1981). A 38,000-year-old archaeological site at Upper Swan, Western Australia. Archaeology in Oceania, 16, 168–172.
Pettitt, P. (2005). The Rise of Modern Humans. In C. Scarre (Ed.), The Human Past (3rd ed., pp. 124–173). Thames and Hudson Ltd.
Potter, B. A., Baichtal, J. F., Beaudoin, A. B., Fehren-Schmitz, L., Haynes, C. V., Holliday, V. T., Holmes, C. E., Ives, J. W., Kelly, R. L., Llamas, B., Malhi, R. S., Miller, D. S., Reich, D., Reuther, J. D., Schiffels, S., & Surovell, T. A. (2018). Current evidence allows multiple models for the peopling of the Americas. Science Advances, 4(8), 1–8.
Roberts, R. G., Jones, R., & Smith, M. A. (1990). Thermoluminescence dating of a 50,000-year-old human occupation site in northern Australia. Nature, 345(6271), 153–156.
Rothhammer, F., & Dillehay, T. (2009). The late pleistocene colonization of South America: an interdisciplinary perspective. The Annals of Human Genetics, 73, 540–549.
Sandweiss, D. H. (1998). Quebrada Jaguay: Early South American Maritime Adaptations. Science, 281(5384), 1830–1832.
Scott, C. (2013). First Footprints – the epic story of the first Australians. Allen & Unwin.
Thoma, A. (1973). New evidence for the polycentric evolution of Homo sapiens. Journal of Human Evolution, 2(6), 529–536.
Turney, C. S. M., Bird, M. I., Fifield, L. K., Roberts, R. G., Smith, M., Dortch, C. E., Grün, R., Lawson, E., Ayliffe, L. K., Miller, G. H., Dortch, J., & Cresswell, R. G. (2001). Early Human Occupation at Devil’s Lair, Southwestern Australia 50,000 Years Ago. Quaternary Research, 55(1), 3–13.
Waguespack, N. M. (2007). Why we’re still arguing about the pleistocene occupation of the Americas. Evolutionary Anthropology, 16, 63–74.
Waters, M., & Stafford, J. T. (2007). Redefining the age of clovis: implications for the peopling of the Americas. Science, 315, 1122–1126.
Williams, T. J., Collins, M. B., Rodrigues, K., Rink, W. J., Velchoff, N., Keen-Zebert, A., Gilmer, A., Frederick, C. D., Ayala, S. J., & Prewitt, E. R. (2018). Evidence of an early projectile point technology in North America at the Gault Site, Texas, USA. Science Advances, 4(7), 1–11.
Alonso, S. & J.A.L. Armour. 2001. A highly variable segment of human subterminal 16p reveals a history of population growth for modern humans outside Africa. Proceedings of the National Academy of Sciences 98: 864–69.
Armitage, S.J., S.A. Jasim, A.E. Marks, A.G. Parker, V.I. Usik & H.-P. Uerpmann. 2011a. The Southern Route “Out of Africa”: Evidence for an Early Expansion of Modern Humans into Arabia. Science 331: 453–56.
Bae, C.J., W. Wang, J. Zhao, S. Huang, F. Tian & G. Shen. 2014. Modern human teeth from Late Pleistocene Luna Cave (Guangxi, China). Quaternary International 354: 169–83.
Bowler, J.M., R. Jones, H. Allen & A.G. Thorne. 1970. Pleistocene human remains from Australia: A living site and human cremation from Lake Mungo, western New South Wales. World Archaeology 2: 39–60.
Cann, R.L., M. Stoneking & A.C. Wilson. 1987. Mitochondrial DNA and human evolution. Nature 325: 31–36.
Caramelli, D. et al. 2003. Evidence for a genetic discontinuity between Neandertals and 24,000-year-old anatomically modern Europeans. Proceedings of the National Academy of Sciences 100: 6593–97.
Cavalli-Sforza, L.L. & M.W. Feldman. 2003. The application of molecular genetic approaches to the study of human evolution. Nature Genetics 33: 266–75.
Cavalli-Sforza, L.L., A. Piazza, P. Menozzi & J. Mountain. 1988. Reconstruction of human evolution: bringing together genetic, archaeological, and linguistic data. Proceedings of the National Academy of Sciences 85: 6002–6.
Clark, J.D. et al. 2003. Stratigraphic, chronological and behavioural contexts of Pleistocene Homo sapiens from Middle Awash, Ethiopia. Nature 423: 747–52.
Coon, C.S. 1962. The Origin of Races. New York: Knopf.
Corny, J. et al. 2017. Dental phenotypic shape variation supports a multiple dispersal model for anatomically modern humans in Southeast Asia. Journal of Human Evolution 112: 41–56.
Darwin C. 1871. The Descent of Man, and Selection in Relation to Sex. London: John Murray.
Deshpande, O., S. Batzoglou, M.W. Feldman & L. Luca Cavalli-Sforza. 2009. A serial founder effect model for human settlement out of Africa. Proceedings of the Royal Society B: Biological Sciences 276: 291–300.
Fadhlaoui-Zid, K., M. Haber, B. Martínez-Cruz, P. Zalloua, A. Benammar Elgaaied & D. Comas. 2013. Genome-Wide and Paternal Diversity Reveal a Recent Origin of Human Populations in North Africa. PLoS ONE 8: 25-26.
Field, J.S. & M.M. Lahr. 2005. Assessment of the Southern Dispersal: GIS-Based Analyses of Potential Routes at Oxygen Isotopic Stage 4. Journal of World Prehistory 19: 1–45.
Finlayson, C. et al. 2006. Late survival of Neanderthals at the southernmost extreme of Europe. Nature 443: 850–53.
Forster, P. & S. Matsumura. 2005. Did Early Humans Go North or South? Science 308: 965–66.
Frumkin, A., O. Bar-Yosef & H.P. Schwarcz. 2011. Possible paleohydrologic and paleoclimatic effects on hominin migration and occupation of the Levantine Middle Paleolithic☆. Journal of Human Evolution 60: 437–51.
Green, R.E. et al. 2008. A Complete Neandertal Mitochondrial Genome Sequence Determined by High-Throughput Sequencing. Cell 134: 416–26.
Groucutt, H.S. et al. 2015a. Human occupation of the Arabian Empty Quarter during MIS 5: evidence from Mundafan Al-Buhayrah, Saudi Arabia. Quaternary Science Reviews 119: 116–35.
—. 2015b. Rethinking the dispersal of Homo sapiens out of Africa. Evolutionary Anthropology: Issues, News, and Reviews 24: 149–64.
Grün, R. et al. 2005. U-series and ESR analyses of bones and teeth relating to the human burials from Skhul. Journal of Human Evolution 49: 316–34.
Gutenkunst, R.N., R.D. Hernandez, S.H. Williamson & C.D. Bustamante. 2009. Inferring the Joint Demographic History of Multiple Populations from Multidimensional SNP Frequency Data. PLoS Genetics 5: 13-18.
Hawks, J. 2001. The Y Chromosome and the Replacement Hypothesis. Science 293: 567–567.
Henn, B.M. et al. 2011. Hunter-gatherer genomic diversity suggests a southern African origin for modern humans. Proceedings of the National Academy of Sciences 108: 5154–62.
Horai, S., K. Hayasaka, R. Kondo, K. Tsugane & N. Takahata. 1995. Recent African origin of modern humans revealed by complete sequences of hominoid mitochondrial DNAs. Proceedings of the National Academy of Sciences 92: 532–36.
Hublin, J.J. 2009. The origin of Neandertals. Proceedings of the National Academy of Sciences 106: 16022–27.
Huerta-Sánchez, E. et al. 2014. Altitude adaptation in Tibetans caused by introgression of Denisovan-like DNA. Nature 512: 194–97.
Huxley, T. 1863. Evidence as to Man’s Place in Nature. London: Williams & Norgate.
Ingman, M., H. Kaessmann, S. Pääbo & U. Gyllensten. 2000. Mitochondrial genome variation and the origin of modern humans. Nature 408: 708–13.
Keinan, A., J.C. Mullikin, N. Patterson & D. Reich. 2009. Accelerated genetic drift on chromosome X during the human dispersal out of Africa. Nature Genetics 41: 66–70.
Kim, H.L., A. Ratan, G.H. Perry, A. Montenegro, W. Miller & S.C. Schuster. 2014. Khoisan hunter-gatherers have been the largest population throughout most of modern-human demographic history. Nature Communications 5: 5692.
Krings, M., A. Stone, R.W. Schmitz, H. Krainitzki, M. Stoneking & S. Pääbo. 1997a. Neandertal DNA Sequences and the Origin of Modern Humans. Cell 90: 19–30.
Kuhn, S.L. 2013. Roots of the Middle Paleolithic in Eurasia. Current Anthropology 54: S255–68.
Lahr, M.M. & R. Foley. 2005. Multiple dispersals and modern human origins. Evolutionary Anthropology: Issues, News, and Reviews 3: 48–60.
Lahr, M.M. & R.A. Foley. 1998. Towards a theory of modern human origins: geography, demography, and diversity in recent human evolution. American journal of physical anthropology Suppl 27: 137–76.
Liu, H., F. Prugnolle, A. Manica & F. Balloux. 2006. A Geographically Explicit Genetic Model of Worldwide Human-Settlement History. The American Journal of Human Genetics 79: 230–37.
Liu, W. et al. 2010. Human remains from Zhirendong, South China, and modern human emergence in East Asia. Proceedings of the National Academy of Sciences 107: 19201–6.
—. 2015. The earliest unequivocally modern humans in southern China. Nature 526: 696–99.
Luis, J.R., D.J. Rowold, M. Regueiro, B. Caeiro, C. Cinnioğlu, C. Roseman, P.A. Underhill, L.L. Cavalli-Sforza & R.J. Herrera. 2004. The Levant versus the Horn of Africa: Evidence for Bidirectional Corridors of Human Migrations. The American Journal of Human Genetics 74: 532–44.
Macaulay, V. et al. 2005. Single, Rapid Coastal Settlement of Asia Revealed by Analysis of Complete Mitochondrial Genomes. Science 308: 1034–36.
McCarthy, R.C. & L. Lucas. 2014. A morphometric re-assessment of BOU-VP-16/1 from Herto, Ethiopia. Journal of Human Evolution 74: 114–17.
McDermott, F., R. Grün, C.B. Stringer & C.J. Hawkesworth. 1993. Mass-spectrometric U-series dates for Israeli Neanderthal/early modern hominid sites. Nature 363: 252–55.
McDougall, I., F.H. Brown & J.G. Fleagle. 2005. Stratigraphic placement and age of modern humans from Kibish, Ethiopia. Nature 433: 733–36.
Meignen, L. 2012. Levantine Perspectives on the Middle to Upper Paleolithic “Transition.” Archaeology, Ethnology and Anthropology of Eurasia 40: 12–21.
Mellars, P. 2006. Going East: New Genetic and Archaeological Perspectives on the Modern Human Colonization of Eurasia. Science 313: 796–800.
Mellars, P., K.C. Gori, M. Carr, P.A. Soares & M.B. Richards. 2013. Genetic and archaeological perspectives on the initial modern human colonization of southern Asia. Proceedings of the National Academy of Sciences 110: 10699–704.
Metspalu, M. et al. 2004. Most of the extant mtDNA boundaries in south and southwest Asia were likely shaped during the initial settlement of Eurasia by anatomically modern humans. BMC genetics 5: 26.
Meyer, M. et al. 2012. A High-Coverage Genome Sequence from an Archaic Denisovan Individual. Science 338: 222–26.
Mijares, A.S. et al. 2010. New evidence for a 67,000-year-old human presence at Callao Cave, Luzon, Philippines. Journal of Human Evolution 59: 123–32.
O’Connell, James.F. & J. Allen. 2012. Forum The Restaurant At The End Of The Universe: Modelling the colonisation of Sahul. Australian Archaeology 74: 5–31.
Oppenheimer, S. 2012a. Out-of-Africa, the peopling of continents and islands: tracing uniparental gene trees across the map. Philosophical Transactions of the Royal Society B: Biological Sciences 367: 770–84.
—. 2012b. A single southern exit of modern humans from Africa: Before or after Toba? Quaternary International 258: 88–99.
Ovchinnikov, I. v., A. Götherström, G.P. Romanova, V.M. Kharitonov, K. Lidén & W. Goodwin. 2000. Molecular analysis of Neanderthal DNA from the northern Caucasus. Nature 404: 490–93.
Pagani, L. et al. 2015. Tracing the Route of Modern Humans out of Africa by Using 225 Human Genome Sequences from Ethiopians and Egyptians. The American Journal of Human Genetics 96: 986–91.
Pemberton, T.J., M. DeGiorgio & N.A. Rosenberg. 2013. Population Structure in a Comprehensive Genomic Data Set on Human Microsatellite Variation. G3 Genes|Genomes|Genetics 3: 891–907.
Petraglia, M. et al. 2007. Middle Paleolithic Assemblages from the Indian Subcontinent Before and After the Toba Super-Eruption. Science 317: 114–16.
Pickrell, J.K. & D. Reich. 2014. Toward a new history and geography of human genes informed by ancient DNA. Trends in Genetics 30: 377–89.
Pope, K.O. & J.E. Terrell. 2007. Environmental setting of human migrations in the circum-Pacific region. Journal of Biogeography 12: 45-56.
Prüfer, K. et al. 2014. The complete genome sequence of a Neanderthal from the Altai Mountains. Nature 505: 43–49.
Prugnolle, F., A. Manica & F. Balloux. 2005. Geography predicts neutral genetic diversity of human populations. Current Biology 15: 159–60.
Qin, P. & M. Stoneking. 2015. Denisovan Ancestry in East Eurasian and Native American Populations. Molecular Biology and Evolution 32: 2665–74.
Ramachandran, S., O. Deshpande, C.C. Roseman, N.A. Rosenberg, M.W. Feldman & L.L. Cavalli-Sforza. 2005. Support from the relationship of genetic and geographic distance in human populations for a serial founder effect originating in Africa. Proceedings of the National Academy of Sciences 102: 15942–47.
Rampino, M.R. & S. Self. 1992. Volcanic winter and accelerated glaciation following the Toba super-eruption. Nature 359: 50–52.
Reed, F.A. & S.A. Tishkoff. 2006. African human diversity, origins and migrations. Current Opinion in Genetics & Development 16: 597–605.
Reich, D. et al. 2010. Genetic history of an archaic hominin group from Denisova Cave in Siberia. Nature 468: 1053–60.
—. 2011. Denisova Admixture and the First Modern Human Dispersals into Southeast Asia and Oceania. The American Journal of Human Genetics 89: 516–28.
Relethford, J.H. 2001. Absence of regional affinities of Neandertal DNA with living humans does not reject multiregional evolution. American Journal of Physical Anthropology 115: 95–98.
Relethford, J.H. 2008. Genetic evidence and the modern human origins debate. Heredity 100: 555–63.
Rito, T., M.B. Richards, V. Fernandes, F. Alshamali, V. Cerny, L. Pereira & P. Soares. 2013. The First Modern Human Dispersals across Africa. PLoS ONE 8: 67-96.
Rosenberg, Karen. 2002. A late Pleistocene human skeleton from Liujiang, China suggests regional population variation in sexual dimorphism in the human pelvis. Variability and Evolution 10: 5–17.
Schlebusch, C.M. et al. 2012. Genomic Variation in Seven Khoe-San Groups Reveals Adaptation and Complex African History. Science 338: 374–79.
Serre, D., A. Langaney, M. Chech, M. Teschler-Nicola, M. Paunovic, P. Mennecier, M. Hofreiter, G. Possnert & S. Pääbo. 2004. No Evidence of Neandertal mtDNA Contribution to Early Modern Humans. PLoS Biology 2: 57-86.
Shea, J.J. 2001. The archaeology of an illusion: the middle-upper paleolithic transition in the Levant, in J.M. le Tenesor, R. Jagher & R. Otte (ed.) The Lower and Middle Paleolithic in the Middle East and Neighbouring Regions.: 169–82. ERAUL 126. Liége: Université de Liége.
Shi, W. et al. 2010. A Worldwide Survey of Human Male Demographic History Based on Y-SNP and Y-STR Data from the HGDP-CEPH Populations. Molecular Biology and Evolution 27: 385–93.
Smith, T.M., P. Tafforeau, D.J. Reid, R. Grün, S. Eggins, M. Boutakiout & J.-J. Hublin. 2007. Earliest evidence of modern human life history in North African early Homo sapiens. Proceedings of the National Academy of Sciences 104: 6128–33.
Soares, P. et al. 2012a. The Expansion of mtDNA Haplogroup L3 within and out of Africa. Molecular Biology and Evolution 29: 915–27.
Stringer, C. 1992. Replacement, continuity and the origin of Homo sapiens, in G. Brauer & F.H. Smith (ed.) Continuity or Replacement? Controversies in Homo sapiens Evolution: 9–24. Rotterdam: Balkema.
Stringer, C. 2000. Coasting out of Africa. Nature 405: 25–27.
—. 2002. Modern human origins: progress and prospects. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences 357: 563–79.
Stringer, C.B. & P. Andrews. 1988. Genetic and fossil evidence for the origin of modern humans. Science (New York, N.Y.) 239: 1263–68.
Takahata, N., S.-H. Lee & Y. Satta. 2001. Testing Multiregionality of Modern Human Origins. Molecular Biology and Evolution 18: 172–83.
Tattersall, I. 2009. Human origins: Out of Africa. Proceedings of the National Academy of Sciences 106: 16018–21.
Templeton, A.R. 2007. Genetics and recent human evolution. Evolution; international journal of organic evolution 61: 1507–19.
Thomson, R., J.K. Pritchard, P. Shen, P.J. Oefner & M.W. Feldman. 2000. Recent common ancestry of human Y chromosomes: Evidence from DNA sequence data. Proceedings of the National Academy of Sciences 97: 7360–65.
Torroni, A., A. Achilli, V. Macaulay, M. Richards & H.-J. Bandelt. 2006. Harvesting the fruit of the human mtDNA tree. Trends in genetics : TIG 22: 339–45.
Underhill, P.A. & T. Kivisild. 2007. Use of Y Chromosome and Mitochondrial DNA Population Structure in Tracing Human Migrations. Annual Review of Genetics 41: 539–64.
Underhill, P.A. et al. 2000. Y chromosome sequence variation and the history of human populations. Nature Genetics 26: 358–61.
Usik, V.I., J.I. Rose, Y.H. Hilbert, P. van Peer & A.E. Marks. 2013. Nubian Complex reduction strategies in Dhofar, southern Oman. Quaternary International 300: 244–66.
Vigilant, L., M. Stoneking, H. Harpending, K. Hawkes & A.C. Wilson. 1991. African Populations and the Evolution of Human Mitochondrial DNA. Science 253: 1503–7.
Walter, R.C. et al. 2000. Early human occupation of the Red Sea coast of Eritrea during the last interglacial. Nature 405: 65–69.
Weidenreich, F. 1940. Some problems dealing with ancient man. American Anthropologist 42: 380–82.
Wu, X.Z. 1981. A well-preserved cranium of an archaic type of early Homo sapiens from Dali, China. Scientia Sinica 24: 530–41.
Ziętkiewicz, E. et al. 2003. Haplotypes in the Dystrophin DNA Segment Point to a Mosaic Origin of Modern Human Diversity. The American Journal of Human Genetics 73: 994–1015.
Arzarello, M., Marcolini, F., Pavia, G., Pavia, M., Petronio, C., Petrucci, M., Rook, L., & Sardella, R. (2007). Evidence of earliest human occurrence in Europe: the site of Pirro Nord (Southern Italy). Naturwissenschaften, 94(2), 107–112.
Berger, L. R., Hawks, J., de Ruiter, D. J., Churchill, S. E., Schmid, P., Delezene, L. K., Kivell, T. L., Garvin, H. M., Williams, S. A., DeSilva, J. M., Skinner, M. M., Musiba, C. M., Cameron, N., Holliday, T. W., Harcourt-Smith, W., Ackermann, R. R., Bastir, M., Bogin, B., Bolter, D., … Zipfel, B. (2015). Homo naledi, a new species of the genus Homo from the Dinaledi Chamber, South Africa. ELife, 4.
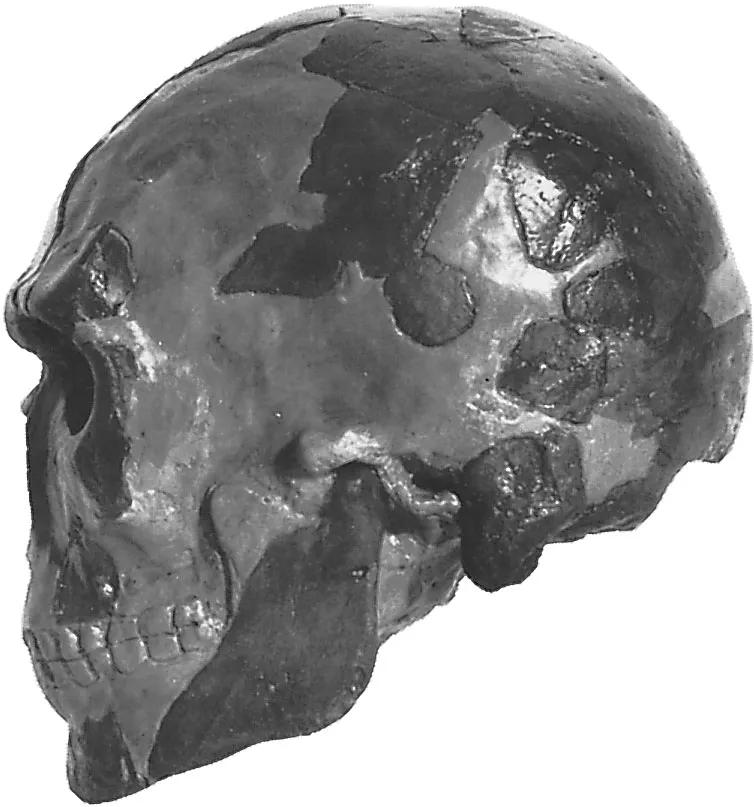
Bermúdez de Castro, J. M., Arsuaga, J. L., Carbonell, E., Rosas, A., Martı́nez, I., & Mosquera, M. (1997). A Hominid from the Lower Pleistocene of Atapuerca, Spain: Possible Ancestor to Neandertals and Modern Humans. Science, 276(5317), 1392–1395.
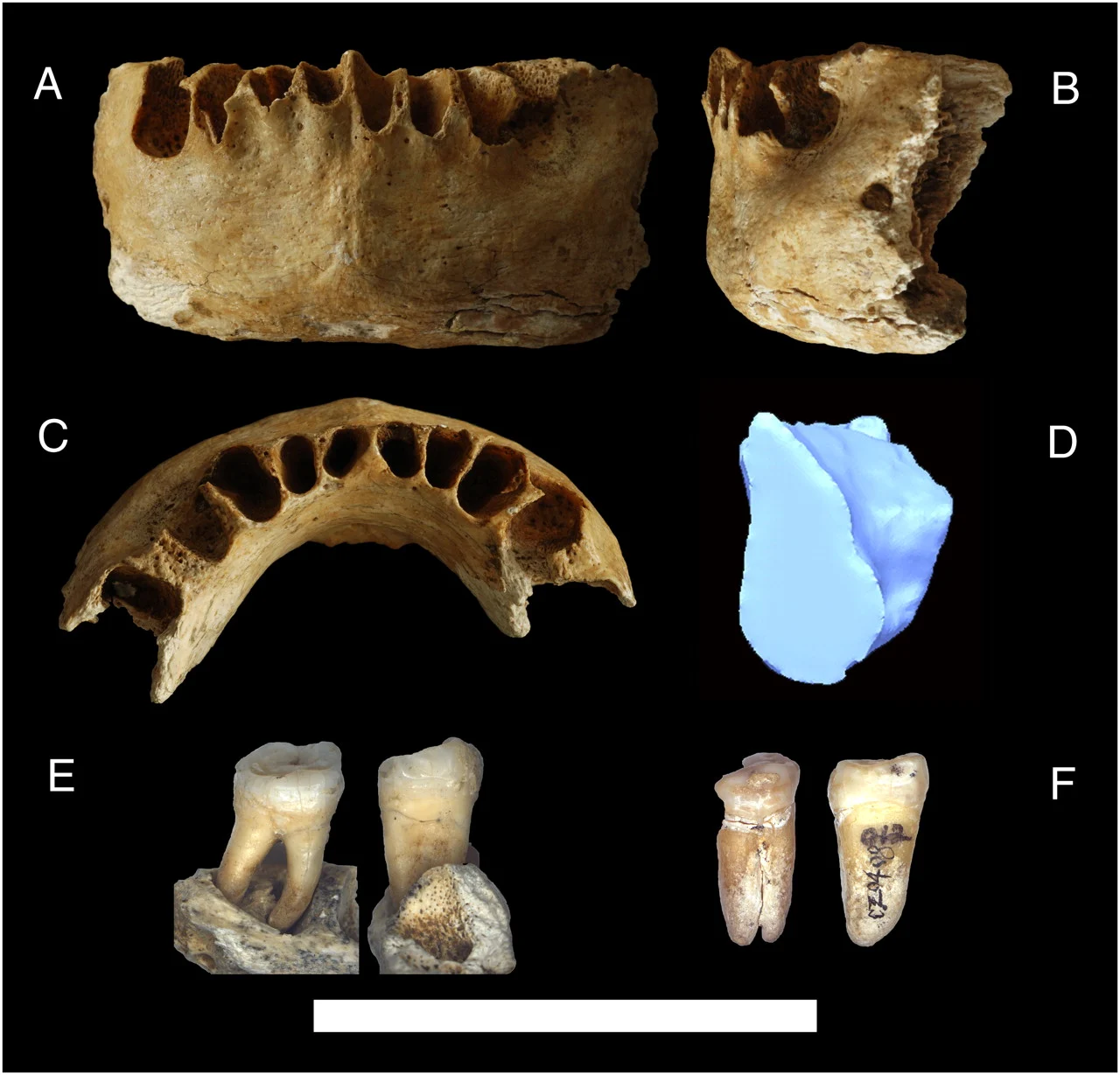
Bermúdez-de-Castro, J.-M., Martinón-Torres, M., Martín-Francés, L., Modesto-Mata, M., Martínez-de-Pinillos, M., García, C., & Carbonell, E. (2017). Homo antecessor : The state of the art eighteen years later. Quaternary International, 433, 22–31.
Bolter, D. R., Hawks, J., Bogin, B., & Cameron, N. (2018). Palaeodemographics of individuals in Dinaledi Chamber using dental remains. South African Journal of Science, 114(1/2).
Brown, S., Higham, T., Slon, V., Pääbo, S., Meyer, M., Douka, K., Brock, F., Comeskey, D., Procopio, N., Shunkov, M., Derevianko, A., & Buckley, M. (2016). Identification of a new hominin bone from Denisova Cave, Siberia using collagen fingerprinting and mitochondrial DNA analysis. Scientific Reports, 6(1), 23559.
Carbonell, E., Bermúdez de Castro, J. M., Parés, J. M., Pérez-González, A., Cuenca-Bescós, G., Ollé, A., Mosquera, M., Huguet, R., van der Made, J., Rosas, A., Sala, R., Vallverdú, J., García, N., Granger, D. E., Martinón-Torres, M., Rodríguez, X. P., Stock, G. M., Vergès, J. M., Allué, E., … Arsuaga, J. L. (2008). The first hominin of Europe. Nature, 452(7186), 465–469.
Carbonell, E., Esteban, M., Nájera, A. M., Mosquera, M., Rodrı´guez, X. P., Ollé, A., Sala, R., Vergès, J. M., Bermu´dez de Castro, J. M., & Ortega, A. I. (1999). The Pleistocene site of Gran Dolina, Sierra de Atapuerca, Spain: a history of the archaeological investigations. Journal of Human Evolution, 37(3–4), 313–324.
Détroit, F., Mijares, A. S., Corny, J., Daver, G., Zanolli, C., Dizon, E., Robles, E., Grün, R., & Piper, P. J. (2019). A new species of Homo from the Late Pleistocene of the Philippines. Nature, 568(7751), 181–186.
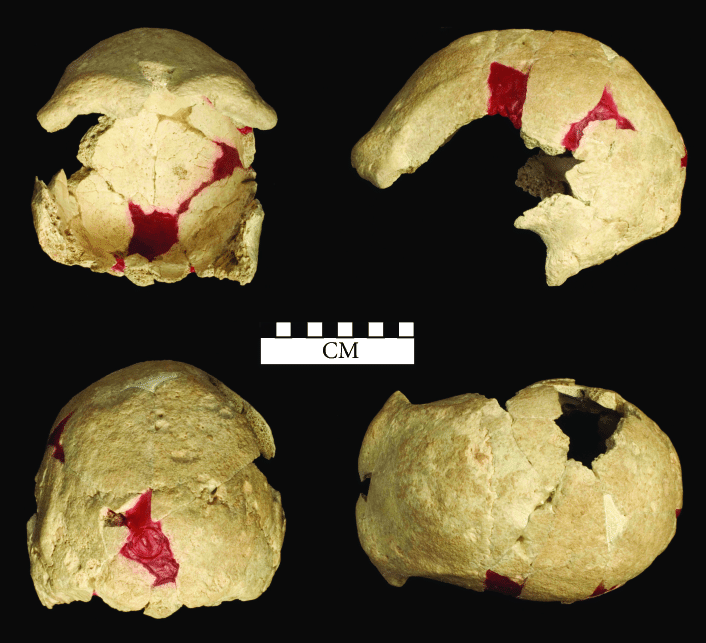
Dirks, P. H., Berger, L. R., Roberts, E. M., Kramers, J. D., Hawks, J., Randolph-Quinney, P. S., Elliott, M., Musiba, C. M., Churchill, S. E., de Ruiter, D. J., Schmid, P., Backwell, L. R., Belyanin, G. A., Boshoff, P., Hunter, K. L., Feuerriegel, E. M., Gurtov, A., Harrison, J. du G., Hunter, R., … Tucker, S. (2015). Geological and taphonomic context for the new hominin species Homo naledi from the Dinaledi Chamber, South Africa. ELife, 4.
Dirks, P. H., Roberts, E. M., Hilbert-Wolf, H., Kramers, J. D., Hawks, J., Dosseto, A., Duval, M., Elliott, M., Evans, M., Grün, R., Hellstrom, J., Herries, A. I., Joannes-Boyau, R., Makhubela, T. v, Placzek, C. J., Robbins, J., Spandler, C., Wiersma, J., Woodhead, J., & Berger, L. R. (2017). The age of Homo naledi and associated sediments in the Rising Star Cave, South Africa. ELife, 6.
Fu, Q., Hajdinjak, M., Moldovan, O. T., Constantin, S., Mallick, S., Skoglund, P., Patterson, N., Rohland, N., Lazaridis, I., Nickel, B., Viola, B., Prüfer, K., Meyer, M., Kelso, J., Reich, D., & Pääbo, S. (2015). An early modern human from Romania with a recent Neanderthal ancestor. Nature, 524(7564), 216–219.
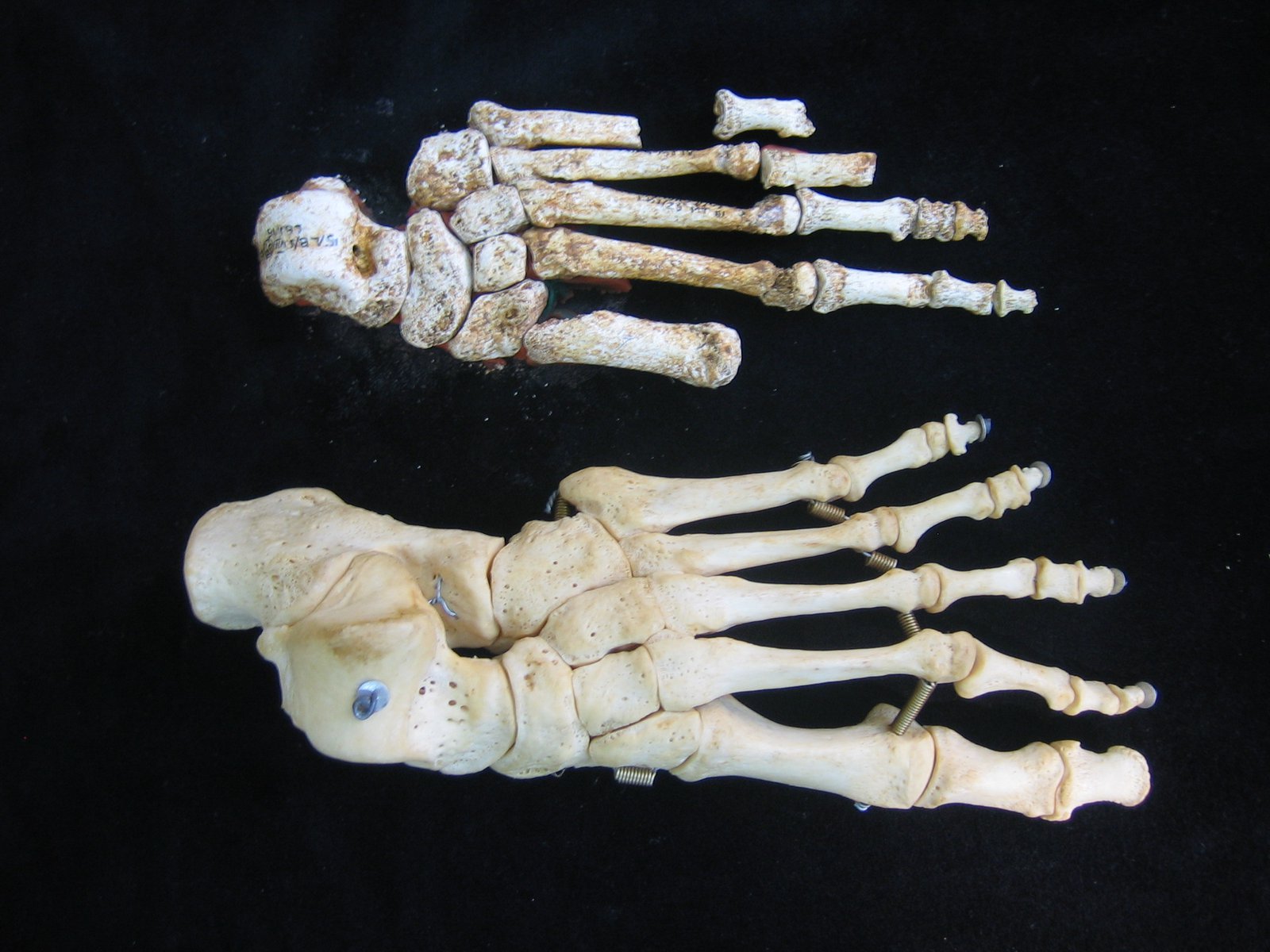
Harcourt-Smith, W. E. H., Throckmorton, Z., Congdon, K. A., Zipfel, B., Deane, A. S., Drapeau, M. S. M., Churchill, S. E., Berger, L. R., & DeSilva, J. M. (2015). The foot of Homo naledi. Nature Communications, 6(1), 8432.
Hawks, J., Elliott, M., Schmid, P., Churchill, S. E., Ruiter, D. J. de, Roberts, E. M., Hilbert-Wolf, H., Garvin, H. M., Williams, S. A., Delezene, L. K., Feuerriegel, E. M., Randolph-Quinney, P., Kivell, T. L., Laird, M. F., Tawane, G., DeSilva, J. M., Bailey, S. E., Brophy, J. K., Meyer, M. R., … Berger, L. R. (2017). New fossil remains of Homo naledi from the Lesedi Chamber, South Africa. ELife, 6.
Kaifu, Y., Kono, R. T., Sutikna, T., Saptomo, E. W., Jatmiko, & Due Awe, R. (2015). Unique Dental Morphology of Homo floresiensis and Its Evolutionary Implications. PLOS ONE, 10(11), e0141614.
Kivell, T. L., Deane, A. S., Tocheri, M. W., Orr, C. M., Schmid, P., Hawks, J., Berger, L. R., & Churchill, S. E. (2015). The hand of Homo naledi. Nature Communications, 6(1), 8431.
Krause, J., Fu, Q., Good, J. M., Viola, B., Shunkov, M. v., Derevianko, A. P., & Pääbo, S. (2010). The complete mitochondrial DNA genome of an unknown hominin from southern Siberia. Nature, 464(7290), 894–897.
López-Polín, L., Bermúdez de Castro, J. M., & Carbonell, E. (2017). The preparation and conservation treatments of the human fossils from Lower Pleistocene unit TD6 (Gran Dolina site, Atapuerca) – The 2003–2009 record. Quaternary International, 433, 251–262.
Mijares, A. S., Détroit, F., Piper, P., Grün, R., Bellwood, P., Aubert, M., Champion, G., Cuevas, N., de Leon, A., & Dizon, E. (2010). New evidence for a 67,000-year-old human presence at Callao Cave, Luzon, Philippines. Journal of Human Evolution, 59(1), 123–132.
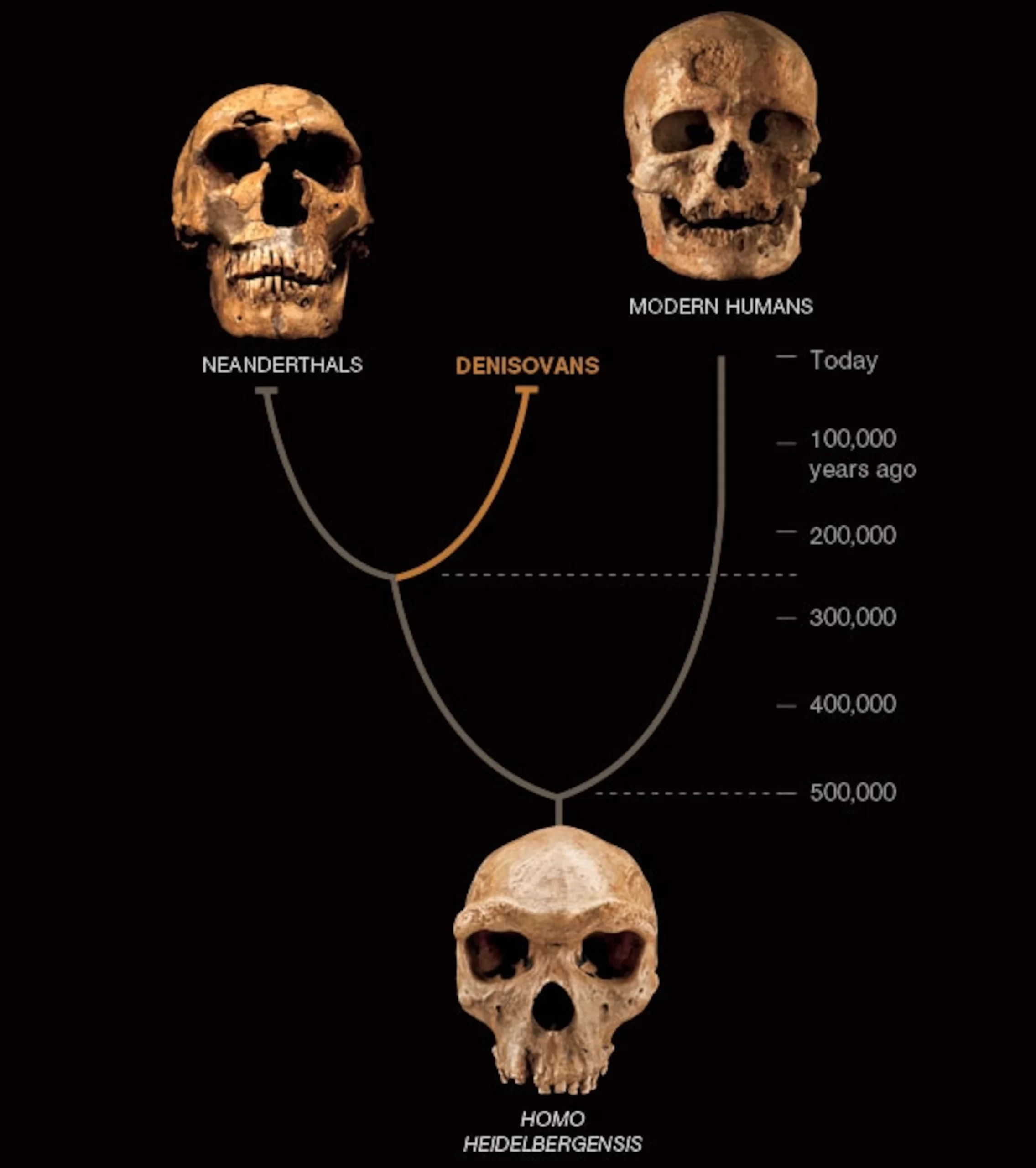
Roksandic, M., Radović, P., Wu, X., & Bae, C. J. (2022). Resolving the “muddle in the middle”: The case for Homo bodoensis sp. nov. Evolutionary Anthropology: Issues, News, and Reviews, 31(1), 20–29.
Rosell, J., Rodríguez, J., & Benito-Calvo, A. (2017). What’s happening now in Atapuerca? Latest research at the Sierra de Atapuerca sites. Quaternary International, 433, 2–4.
Slon, V., Mafessoni, F., Vernot, B., de Filippo, C., Grote, S., Viola, B., Hajdinjak, M., Peyrégne, S., Nagel, S., Brown, S., Douka, K., Higham, T., Kozlikin, M. B., Shunkov, M. v., Derevianko, A. P., Kelso, J., Meyer, M., Prüfer, K., & Pääbo, S. (2018). The genome of the offspring of a Neanderthal mother and a Denisovan father. Nature, 561(7721), 113–116.
Warren, M. (2018). Mum’s a Neanderthal, Dad’s a Denisovan: First discovery of an ancient-human hybrid. Nature, 560(7719), 417–418.
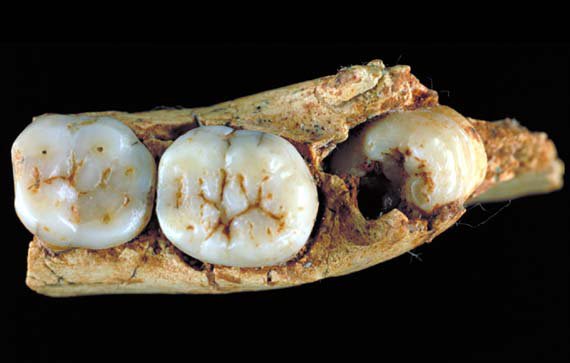
Welker, F., Ramos-Madrigal, J., Gutenbrunner, P., Mackie, M., Tiwary, S., Rakownikow Jersie-Christensen, R., Chiva, C., Dickinson, M. R., Kuhlwilm, M., de Manuel, M., Gelabert, P., Martinón-Torres, M., Margvelashvili, A., Arsuaga, J. L., Carbonell, E., Marques-Bonet, T., Penkman, K., Sabidó, E., Cox, J., … Cappellini, E. (2020). The dental proteome of Homo antecessor. Nature, 580(7802), 235–238.













 සම්මාණයෙන් පිදුම් ලබයි")