{kind=link}
කායච්ඡේද විද්යාත්මකව නූතන මානවයා අප්රිකාවෙන් පිට වී යාම පිළිබදව වූ පළමු සංක්රමණය පිළිබද සිදුවන විද්යාත්මක පර්යේෂණ හා විදග්ධ කතිකාවත වර්තමානය වන විට ද ලොව පුරා පුරවිද්යාඥයින්, මානව විද්යාඥයින් හා පුළුල් පරාසයක විෂය ප්රාමාණිකයින් අතර විශාල උනන්දුවක් ඇති කර ඇත. පවතින ෆොසිල, පුරාවිද්යාත්මක සහ පුරාදේශගුණික දත්ත විෂයානුබද්ධව බොහෝ උපකල්පන ඉදිරිපත් කරන අතර, ඊට අමතර ව, ජානමය වශයෙන් ප්රවේණිගත කිරීම සහ කාලනිර්ණය කිරීමේ ශිල්පීය ක්රම සහ ජාන සාම්පල ලබා ගැනීමේ හැකියාව වැඩි වීමත් සමඟ, වසර මිලියන ගණනක් පැරණි ඉතිහාසයට අදාළ න්යාය පරීක්ෂා කිරීම සඳහා අවශ්ය තාක්ෂණික අංග වැඩිදියුණු වෙමින් පවතී. මෙම ලිපිය මගින්, ආදි මානවයා අප්රිකාවෙන් පිටව ගියේ කෙසේද සහ කවදාද, විසිර යාමේ වාර කීයක් තිබේද සහ භූගෝලීය මාර්ග මොනවාද යන්න පිළිබඳව වන අඛණ්ඩ විවාද පිලිබද අවධානය යොමු කෙරේ. එහි දී ප්රධාන පුරාවිද්යාත්මක සොයාගැනීම් කිහිපයක් සමඟ වත්මන් ජාන විද්යාත්මක පර්යේෂණ භාවිතා කරමින්, ඒවායේ වලංගුභාවය හා අනාගත පර්යේෂණ අවස්ථා පිළිබද අවසන් සාකච්ඡාව මෙහෙය වේ.
අප්රිකානු මහද්වීපයේ සිදුවූ මානව සම්භවය 19 වැනි සියවසේදී චාල්ස් ඩාවින් විසින් ප්රසිද්ධ ලෙස යෝජනා කරන ලදී (Darwin 1871). අප්රිකාවේ චිම්පන්සින් සහ ගෝරිල්ලන් සිටීම සහ නූතන මිනිසුන් සහ වානරයන් පොදු මුතුන් මිත්තෙකු බව හක්ස්ලිගේ සංසන්දනාත්මක ව්යුහ විද්යා අධ්යයනයන් මත පදනම්ව (Huxley 1863), ඩාවින් නූතන මිනිසුන්ගේ මුතුන් මිත්තන් අප්රිකානු භූමියේ සම්භවය විණි යැයි තර්ක කළේය. වර්තමානයේ, වසර 150කට වැඩි කාලයකින් පසුව, පුරාවිද්යාත්මක සාක්ෂි සමඟින් ජානමය වශයෙන් ප්රවේණිගත කිරීම සහ අනුක්රමණය කිරීමේ ශිල්පීය ක්රමවල වැඩිදියුණු වීමත් සමඟ ජාන විද්යාව, තවත් බොහෝ සංකීර්ණතා ඉස්මතු කරන අතරම, පළමු නූතන මානවයාගේ අප්රිකානු සම්භවය සැලකිය යුතු ලෙස ශක්තිමත් අන්දමින් තහවුරු කරයි. විශේෂයෙන්ම, මෙම ප්රවේණික ක්රමවල අඛණ්ඩ දියුණුවත් සමඟ සහ නව ජෛව තොරතුරු ප්රවේශ (Bioinformatics Tools) සමඟ සහජීවන සම්බන්ධ කිරීමක් (symbiotic coupling) හරහා, පර්යේෂකයින් මානව මූලාරම්භය පිළිබඳ ප්රශ්නවලට, පෙර පර්යේෂණවලින් අනාවරණය වූ තොරතුරුවලට සියුම් විභේදනයක් සහිත පිළිතුරු දීමට ජානමය තොරතුරු භාවිතා කිරීමට උත්සාහ කරයි. ඒ අනුව අපේ මුතුන් මිත්තන් මුලින්ම අප්රිකාව හැර ගියේ කෙසේද සහ කවදාද? විසරණ වාර කිහිපයක් හෝ තනි එකක් තිබුණාද? ඔවුන් ගමන් ගත් භූගෝලීය මාර්ග මොනවාද? තවමත් විවාදාපන්න ලෙස, දිනෙන් දින නව්ය වන තාක්ෂණික කාරණා ඔස්සේ වැඩිදියුණු වෙමින් පවතී.
මෙම මාතෘකාව සියුම් ව ගලායන පරිදි කරුණු සම්පිණ්ඩනය කිරීම අපහසු කාර්යයක් වන්නේ, විෂය කරුණුවල ඇති ඝට්ටනය, සංකීර්ණත්වය හා දිනෙන් දින ෆොසිල වාර්තාවේ සිදුවන යාවත්කාලීන කිරීම් හමුවේ එකතුවන නව දැනුම් කරුණු ගලපා ගැනීමට යම් කාලයක් අවශ්ය වන බැවිනි. බොහෝ විට එම කාලය මාස කීපයක සිට වසර කීපයක් දක්වා දිගු විය හැකි ය. එම නිසා කියවා ගැනීමේ හා තේරුම් ගැනීමේ පහසුව උදෙසා මෙතෙක් විද්වතුන් විසින් මානවයා අප්රිකාවෙන් පිට වී යාම පිළිබද පර්යේෂණ තේමා කීපයක් යටතේ ගොනු කර ඇත. නමුත්, පිළිගත යුතු යථාර්තය වන්නේ ඒවායින් එකක් පමණක් කිසිවිටෙකත් සත්යය නොවන බව ය. විටෙක සියල්ලම සත්යය විය හැකි ය, නමුත්, නිශ්චිතවම කායච්ඡේද විද්යාත්මකව ‘නූතන මානවයා මෙම කාලයේ දී, මෙම භුගෝලීය ප්රදේශය භාවිතා කරමින් පමණක් අප්රිකාවෙන් පිටවී ගොස් නව ලෝකය ජනාවාස කළේය’ නම් පරම සත්යයක් විද්යා ලෝකය තුළ දිරිමත් නොකරයි. ඒ අනුව වර්තමානයේ දී පවා උණුසුම් ලෙස විදග්ධ කතිකාවත මෙහෙයවෙන නූතන මානවයා අප්රිකාවෙන් පිට වී යාමේ සංසිද්ධියට අදාල න්යායාත්මක ආකෘති කීපයක් මෙම ලිපියේ ඉදිරි කොටස් මගින් පැහැදිලි කෙරේ.
Candelabra හා ප්රතිස්ථාපන උපකල්පනය (The Replacement Hypothesis)
20 වැනි සියවසේ දෙවන භාගය පුරා මානව විද්යාත්මක කතිකාවතෙහි වැඩි ප්රමාණයක් ආධිපත්යය දැරූ විවාදය විසින්, කායච්ඡේද විද්යාත්මකව නවීන මානවයා (Anatomically Modern Human) ලෙස අප හඳුන්වනු ලබන නූතන හෝමෝ සේපියන්ස් බවට පරිණාමය වූයේ කොතැනද සහ කවදාද යන්න පිළිබඳව අවධානය යොමු කළේය. ප්රධාන උපකල්පන දෙකක් එම කතිකාවෙහි ආධිපත්යය දැරීය: එනම්, බහු කලාපීය උපකල්පනය (Multiregional Hypothesis) සහ ප්රතිස්ථාපන උපකල්පනය (The Replacement Hypothesis) වශයෙනි. මෙකී මුල්කාලීන බහු කලාපීය ආකෘතිය මානව විද්යාඥ Weidenreich විසින් 1946 දී යෝජනා කරන ලද අතර ප්ලයිස්ටොසීනය පුරාවට ලෝකයේ විවිධ ප්රදේශවල ජීවත් වන හෝමෝ ඉරෙක්ටස්ගේ උප ජනගහන අතර සැලකිය යුතු ජාන ප්රවාහයක්, නූතන හෝමෝ සේපියන්වරු බවට පත් වීමට අනුග්රහය දැක්වූ බව එහි මූලික හරය වේ (Weidenreich 1940). නමුත් මෙහිදී සැලකිය යුතු කරුණක් වන්නේ බහු කලාපීය ආකෘතිය බොහෝ විට Candelabra ආකෘතියට සමානව භාවිතා කර ඇති බව ය, (Templeton 2007) එම ආකෘතිය පළමු වරට 1962 දී කූන් විසින් යෝජනා කරන ලදී (Coon 1962). මෙම ආකෘතිය යටතේ ස්වාධීනව පරිණාමය වූ ව්යුහ විද්යාත්මක නවීන ලක්ෂණ සහිත මානව ස්වරූපය පසුගිය වසර මිලියන 1 තුළ ලොව පුරා විවිධ කාලවලදී සහ ස්ථානවලදී ස්වයංක්රීයව පැන නැඟී ඇත (Coon 1962), එබැවින් නූතන අප්රිකානු නොවන ජනගහනය එක් එක් මූලික වශයෙන් මෙම හෝමෝ විශේෂවල වෙනම පරිණාමයන්ගෙන් පැවත එන්නකි. මීට පරිබාහිරව පවතින, බහු කලාපීය ආකෘතිය, කායච්ඡේද විද්යාත්මකව නූතන මානවයාගේ ස්වාධීන හා සමාන්තර පරිණාමය ප්රතික්ෂේප කරයි.

බහු කලාපීය සහ Candelabra උපකල්පනවලට සහාය දක්වන ප්රධාන ෆොසිල සාක්ෂිය වූයේ චීනයේ ඩාලි මිනිසා (Dali Man) සොයා ගැනීමයි (Wu 1981). බහු කලාපීයවාදීන් හෝ Candelabra ආකෘතික ආධාරකරුවන් සඳහා, එය පෞරාණික සහ නවීන ලක්ෂණ මිශ්ර, මුල් හා නූතන හොමිනින් අතර මධ්ය පරිණාමීය ලක්ෂණ ප්රකට වූ සාක්ෂි විය. එනම්, මෙම ෆොසිල දුර්වල සංරක්ෂණය වී ඇතත්, සමහර විද්වතුන් මෙම ව්යුහ විද්යාත්මක ලක්ෂණ ලොව පුරා සිටින අනෙකුත් හෝමෝවරුන් විසින් අඩුවැඩි වශයෙන් උරුමකරගත් බව යෝජනා කර ඇති අතර, එබැවින් එය ආසියාවට ආවේණික නොවන බව කියයි (Stringer & Andrews 1988; Stringer 1992). තවදුරටත් ඔවුන් අප්රිකානු මහාද්වීපයෙන් පිටත ජානමය උප්පත්ති ස්ථාන කිහිපයක මූලාරම්භය අනුමාන කර ඇත. උදාහරණ ලෙස මානව Dystrophin ජානයේ පැරණිතම හැප්ලෝටයිපය අප්රිකානුවන් තුළ නොමැති බව සොයා ගන්නා ලදී (Ziętkiewicz et al. 2003), එහි පශ්චාත් විශ්ලේෂණ කටයුතුවල දී අනාවරණය වූයේ Candelabra ආකෘතියට සහය ලබා දෙනවාට වඩා නියැන්ඩර්තාල්වරුන්ගෙන් අනුවර්තනය වීමේ ප්රතිඵලයක් ලෙසිනි.
Candelabra ආකෘතියට එදිරිවන සාධක පාෂාණ විද්යාත්මක (paleontological) සහ ජාන අධ්යයනයන් (genetic studies) තුළින් අනාවරණය වී ඇත. ප්රතිස්ථාපනය, හෝ අප්රිකාවෙන් පිට වී යාමේ න්යාය (Out of Africa), ආකෘතිය යෝජනා කරන්නේ අතීත මානවයින්ගේ සිට අප්රිකාවේ කායච්ඡේද විද්යාත්මකව නූතන මානවයා දක්වා වූ ගමන, හුදකලා සහ සාපේක්ෂව මෑත සංක්රමණයක් වන අතර, පසුව ලොක්යේ අනෙකුත් ප්රදේශවල ඒ වන විටත් ජනාවාසගතව සිටි ජනගහනය රළු ලෙස ප්රතිස්ථාපනය කරමින් ලෝකයේ සෙසු ප්රදේශවලට පසුව සංක්රමණය වීම එම ආකෘතියයේ මූලික හරයයි (Stringer & Andrews 1988; Ingman et al. 2000; Stringer 2002; Cavalli-Sforza & Feldman 2003; Relethford 2008; Tattersall 2009). මෙම ආකෘතිය යටතේ සේපියන්වරු හැර අනෙකුත් මානව වර්ගවල වදවී යාම දිරිමත් කරන අතර, සමකාලීන සේපියන් ගහනයේ බොහෝ ජානමය විවිධත්වය අදින් වසර 55,000 හා 200,000 අතර කාලය තුළ දී පැවත එන බවත්, එම කාලය කායච්ඡේද විද්යාත්මකව නූතන මානවයා තනි හෝ බහු කණ්ඩායම් ප්රවේශයකින් අප්රිකාවෙන් පිට වී ගිය බවත් බොහෝ පර්යේෂකයින්ගේ අදහසයි.
අප්රිකාවෙන් පිටවී යාමේ න්යායට අනුරූප වන පළමු ජානමය සාක්ෂි සපයන ලද්දේ මයිටොකොන්ඩ්රියල් DNA හෙවත් මව්පාර්ශවයේ ජාන (Mitochondrial DNA) හි ජානවිද්යාත්මක ගස (Phylogenetic Tree) අධ්යයනය කිරීමෙනි, එය මානව මාතෘජාන සංචිතයේ මූලාශ්රය ලෙස අප්රිකාව හඳුනාගෙන ඇත (Cann et al. 1987; Vigilant et al. 1991; Horai et al. 1995; Tattersall 2009). එමගින් තවදුරටත් අප්රිකාවෙන් පිටත ඇති සියලුම මාතෘජාන සංචිතයේ (mtDNA haplogroups) දකුණු ආසියාවේ අදින් වසර 60,000- 65,000 ක් දක්වා ඇති M හෝ N ජාන සංචිතවලට ආරෝපණය කළ හැකි බව පෙන්වා දෙන ලද අතර (Macaulay et al. 2005), එය අදින් වසර 80,000 සිට පමණ නැගෙනහිර අප්රිකාවේ ඇති වූ බවට උපකල්පනය කරන ලද L3 ජාන සංචිතයෙන් පැවත එන බව සැලකේ (Metspalu et al. 2004). මානවයාගේ අප්රිකානු සම්භවය තවදුරටත් මාතෘජාන (mtDNA) (Ovchinnikov et al. 2000; Relethford 2001; Caramelli et al. 2003), Y chromosome (Thomson et al. 2000; Underhill et al. 2000; Hawks 2001) හා autosomal regions (Alonso & Armour 2001; Takahata et al. 2001) තුළින් තහවුරු වී ඇත. තවදුරටත් පුළුල් ජානමය දත්ත විශ්ලේෂනයේ දී තහවුරු වී ඇත්තේ, නැගෙනහිර හෝ දකුණු අප්රිකාවේ සිට භූගෝලීය දුර ප්රමාණයේ ශ්රිතයක් ලෙස ගත්විට එහි ප්රවේණිගත ජාන විවිධත්වය අඩුවන බව ය, එනම් විෂමජාතීයතාවයේ ආසන්න වශයෙන් සිදුවන රේඛීය අඩුවීමක් සහ සම්බන්ධතා අසමතුලිතතාවය අප්රිකාවෙන් පිටවී යාමේ න්යායට අනුකූල වන බව ය (Prugnolle et al. 2005; Ramachandran et al. 2005).
නූතන මානවයා හා ඔවුන්ගේ දකුණු, නැගෙනහිර හා උතුරු අප්රිකානු සම්භවය
කෙසේ වෙතත්, කායච්ඡේද විද්යාත්මක නූතන මානවයාගේ අප්රිකානු සම්භවය දැන් බොහෝ දුරට පිළිගෙන ඇතත්, ව්යුහ විද්යාත්මකව නවීන ස්වරූපය මුලින්ම නැඟෙනහිර, දකුණ හෝ උතුරු අප්රිකාවේ ඇති වූයේද යන්න පිළිබඳව විවාදය වර්තමානය දක්වාම පවතී.
දකුණු අප්රිකානු සම්භවය
නැගෙනහිර අප්රිකානු සම්භවයක් සඳහා සහය සපයනු ලබන්නේ ඉතියෝපියාවේදී මේ දක්වා පැරණිතම (අදින් වසර 190,000 හා 200,000) අවිවාදිත නවීන මානව ෆොසිල සොයාගැනීමෙනි: Omo I ක්ෂේත්රයේ Kibish ප්රදේශයේ තිබී වර්ෂ 1967 දී (Clark et al. 2003; McDougall et al. 2005) හා Herto ෆොසිල (McCarthy & Lucas 2014) (අදින් වසර 160,000 හා 154,000 දක්වා පැරණි) රිචඩ් ලීකී විසින් විසින් ප්රථම වරට සොයා ගන්නා ලදී.
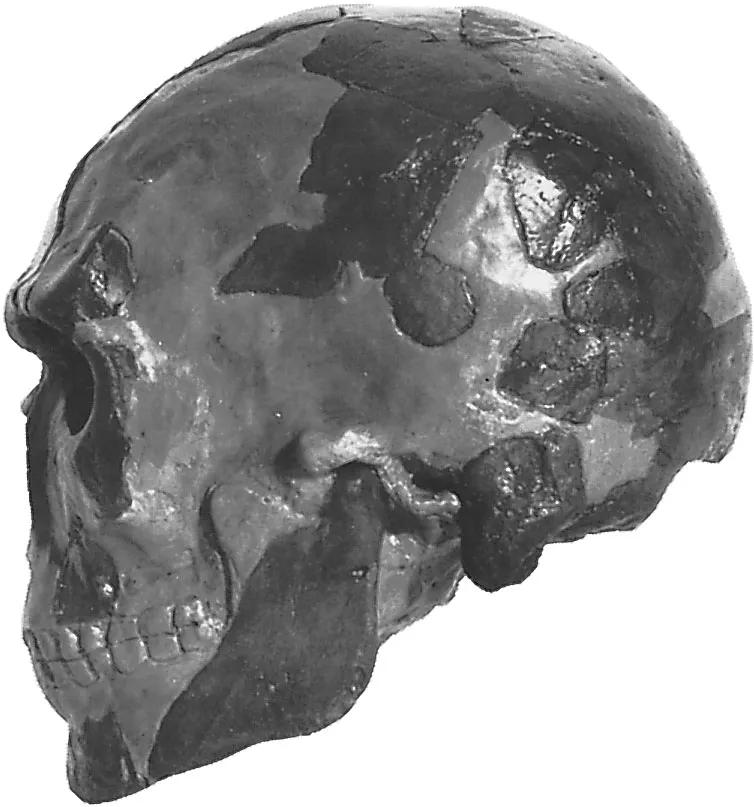

දක්ෂිණාංශික සම්භවයක් සඳහා සාක්ෂි බොහෝ දුරට ලබා දී ඇත්තේ පුරාවිද්යාත්මක අධ්යයනයන්ට වඩා ජානමය අධ්යයන මගිනි. හෙන් ඇතුළු පර්යේෂණ කණ්ඩායම විසින් උප සහරා අප්රිකාවේ දැනට සිටින දඩයම්කරුවන්ගෙන් ප්රවේණි විශ්ලේෂණය කරන ලදී: මධ්යම අප්රිකාවේ පිග්මි, නැගෙනහිර අප්රිකාවේ ටැන්සානියාවේ ආදිවාසීන් (Hazda සහ Sandawe) සහ දකුණු අප්රිකාවේ Khomani Bushmen ඔවුන් තම අධ්යනය සදහා තෝරාගන්නා ලදී (Henn et al. 2011). එහි ප්රතිඵල පෙන්වා දෙන පරිදි ඔවුන්ගේ LD (Linkage Disequilibrium) සහ Heterozygosity රටා තුළින් දකුණු අප්රිකානු දඩයම්කරුවන් සියලු මානව ජනගහනයෙන් වඩාත්ම ජානමය වශයෙන් විවිධ වූවන් අතර වන අතර, එය මානවයා අප්රිකාවෙන් පිටවී යාමේ න්යාය සදහා ද පිටිවහලක් ලෙස සැලකේ. දකුණු අප්රිකානු ආදිවාසී ජනසමාජ වල Linkage disequilibrium කේන්ද්රකරගත් තවත් පර්යේෂණ තුළින් උක්ත පර්යෙෂණයේ සොයාගැනීම් තහවුරු කරඇති අතර, අප්රිකාවේ අනෙකුත් ප්රදේශවල එම අගය සාපේක්ෂව අඩුඅගයක් ගන්නා බව ද සන්සන්දනාත්මක ව ඉදිරිපත් කර ඇත (Schlebusch et al. 2012).
වඩාත් මෑතකාලීන අධ්යයනයක් මගින් ලෝකයේ වෙනත් ආදිවාසී කණ්ඩායම් හා පුරාණ පාරිසරික දත්ත උපයෝගී කරගෙන පුරාණ ජනගහන ප්රමාණය පිළිබඳ ඇස්තමේන්තු ඒකාබද්ධව විශ්ලේෂණය කරන ලදී (Kim et al. 2014). එමෙන් ම මාතෘජාන සංචිත ව්යාප්තිය පිළිබද සිදුකරන ලද පර්යේෂණයක් තුළින් නවීන මාතෘජාන සංචිත ආරම්භ වූ පූර්වජයා දකුණු අප්රිකානු කණ්ඩායම් වලට බෙදීමට පෙර මධ්යම අප්රිකාවේ දී සම්භවය වූ බව අනාවරණය කරන ලදී (Rito et al. 2013).

උතුරු අප්රිකානු සම්භවය
මෑතක් වන තුරුම, අප්රිකාවෙන් පිටවී යාම පිලිබද ආකෘතියේ විභව මූලාරම්භක ලක්ෂ්යයක් ලෙස උතුරු අප්රිකාව කෙරෙහි එතරම් අවධානයක් යොමු කර නැත, නමුත් මොරොක්කෝවේ Jebel Irhoud හි අනාවරණය කරගත් සාධක අදින් වසර 160,000 ක් දක්වා දිව යයි (Smith et al. 2007). වරින් වර සංශෝධනය කරන ලද ජාන ගසෙහි (Phylogenetic tree) පැරණිතම ජාන සමූහ, මධ්යම සහ වයඹදිග අප්රිකාවේ මුල් බැස ඇති බව සොයා ගත් අතර, එමගින් මෙම කලාපය කෙරෙහි අවධානය වැඩි වශයෙන් යොමු විය. උතුරු අප්රිකාව, මැදපෙරදිග හා යුරෝපයේ ව්යාප්ත වී ඇති Y chromosome හා වෙනක් ජාන විශේෂිත සාධක පර්යේෂණයට ලක් කිරීම මගින් නූතන මානවයාගේ උතුරු අප්රිකානු සම්භවය සනාථ කරයි (Fadhlaoui-Zid et al. 2013).

අප්රිකාවෙන් පිට විය හැකි මාර්ග
නූතන මානවයා අප්රිකාවෙන් පිටවීම සම්බන්ධයෙන් වඩාත් කුතුහලය දනවන ප්රශ්නයක් වන්නේ ඔවුන් කුමන භූගෝලීය මාර්ගයක් භාවිත කළේ ද යන්නයි. පිළිගත් මතය නම්, නූතන මානවයන් එක් විසුරුමකින් අප්රිකාවෙන් පිටව ගියේ නම්, එකල පැවතිය හැකි මාර්ග දෙකක් පිළිබද තොරතුරු දැක්විය හැකි හැකි ය: එනම්, උතුරු මාර්ගය ලෙස, ඊජිප්තුව සහ සීනායි හරහාත්, දකුණු මාර්ගය ලෙස, ඉතියෝපියාව හරහා, Bab el Mandeb සමුද්ර සන්ධිය සහ අරාබි අර්ධද්වීපයත් යන්න ය. නමුත්, වර්තමානය දක්වා ම මේ ප්රශ්නය විශ්වාසයෙන් යුතුව විසඳා ගැනීමට පුරාවිද්යාත්මක හෝ ජානමය සාක්ෂිවලට මෙතෙක් හැකි වී නැත.
උතුරු දොරටුව
අප්රිකාවෙන් පිටත දී හමුවන නූතන සේපියන් ෆොසිල වන්නේ Skhul සහ Qafzeh, Levant හි හමුවන ෆොසිල වන අතර, ඒවා සියල්ල අදින් වසර 120,000 හෝ 100,000 – 90,000 දක්වා පැරණි ය (Grün et al. 2005). මෙකී ෆොසිල සිනායි අර්ධද්වීපය හරහා ලෙවන්ට් දක්වා ගමන් කරන, ආසන්න වශයෙන් අදින් වසර 120,000 පැරණි මුල් සේපියන්වරුන්ට අයත් පිටවීමක් නියෝජනය කරන බවට යෝජනා වී ඇත. මෙම කලාපයෙන් හමුවූ මීළඟ මානව ෆොසිල අතර ලෙවන්ට්හි මුල් සේපියන් ජනාවාස, ස්තරානුකූලව සැලකිය යුතු පරතරයක් පෙන්නුම් කරමින් අදින් වසර 55,000 -59,000 ක් පමණ කාල නිර්ණය කරන ලද Manot1 හිස් කබල අනාවරණය වේ. මෙය, දේශගුණික වාර්තා සමඟ ඒකාබද්ධව, ගෝලීය ග්ලැසියර කාලපරිච්ඡේදය අදින් වසර 90,000- 60,000 ක් අතර කාලයේ දී පළමු මිනිසුන් ලෙවන්ට් මිනිසුන් එහි ජීවත් වුවද පැවති දේශගුණික විපර්යාස තුළින් ඔවුන්ගේ අවශේෂ විනාශ වන්නට ඇති බව විද්වතුන්ගේ මතය යි (Pope & Terrell 2007). අනෙකුත් කතුවරුන් අවධාරණය කරන්නේ අදින් වසර 90,000කට පෙර එම සේපියන්ස්වරු ලෙවන්ට් හැර ප්රදේශය හැරගිය බව ය. තවදුරටත් අරාබි අර්ධද්වීපයෙන් පුරාවිද්යාත්මක ද්රව්ය, මූලික වශයෙන් අදින් වසර 100,000 – 90,000 දක්වා පැරණි ගල් ආයුධ සහ පුරාකෘති එකතූන් මගින් ඇඟවෙන්නේ එම ප්රදේශයේ මුල් ජනාවාස පුළුල් ලෙස පැතිරී තිබූ බවත්, Skhul සහ Qafzeh ෆොසිල වාර්තා මගින් ලෙවාන්ට් ප්රදේශයේ සිදුවූ අසාර්ථක නික්මයෑමක් නියෝජනය කරන බවත් ය (Groucutt et al. 2015a).

පුරාවිද්යාත්මක හා දේශගුණික වාර්තා වලින් ලැබෙන සාක්ෂි වලට අමතරව, ජාන අධ්යයනයන්ද උතුරු මාර්ගයක් සඳහා යම් සහයෝගයක් ලබා දී ඇත. microsatellite loci 10ක් හා binary markers 45ක් සමග Y chromosome ජාන සංචිතයේ ව්යාප්තිය නිරීක්ෂණය කළ පර්යේෂණයකින් අප්රිකාව සහ යුරේසියාව අතර ප්රාථමික සංක්රමණික චලනයන් සඳහා ලෙවන්ට් ප්රදේශය වඩාත් ආධාරක වූ මාර්ගය බවට නිගමනය කර ඇත (Luis et al. 2004). තවත් පර්යේෂණයකින් ඉතියෝපියානු ජනවාර්ගික කණ්ඩායම් පහකින් (Amhara, Oromo, Ethiopian Somali, Welayta, සහ Gumuz) ඊජිප්තුවරුන් 100 දෙනෙකුගේ සහ පුද්ගලයන් 125 දෙනෙකුගේ ප්රවේණි අනුපිළිවෙලක් සකස් කරන ලදී. ඒ අනුව පසුගිය වසර 4000 තුළ අප්රිකානු ජාන විශ්ලේෂණය, නවීන අප්රිකානු නොවන ජාන, ඉතියෝපියානු ජානවලට වඩා ඊජිප්තු ජාන වලට සමාන බව ඔවුන් පෙන්වා දුන් අතර, එමඟින් ඔවුන්ගේ කාර්යක්ෂමතාව උපකල්පනය කරමින් අප්රිකානු සංක්රමණයේ වැඩි ඉඩක් ඇති මාර්ගය ඊජිප්තුව හරහා වැටී ඇති බව යෝජනා කරයි (Pagani et al. 2015). කෙසේ වෙතත්, නූතන ජාන විශ්ලේෂණ අධ්යයනයන්හි එක් සීමාවක් නම්, ජනගහන ප්රතිස්ථාපනය, සංක්රමණය, මිශ්රණය සහ ප්ලාවිතය වැනි සාධක හේතුවෙන් පවතින ජනගහනය අතීත ජනගහනයේ ප්රමිතිගත නියෝජිතයන් නොවිය හැකි බවයි.
දක්ෂිණ දොරටුව
උතුරු දොරටුවෙන් පරිබාහිර ව, මාතෘජාන අධ්යයන සම්ප්රදායිකව රතු මුහුදේ Bab el Mandeb සමුද්ර සන්ධිය හරහා වූ බවට වන සාක්ෂි මගින් නූතන සේපියන් අප්රිකාවෙන් පිටවී යාම දක්ෂිණාංශික මාර්ගයකින් සිදු වූ බවට අදහස් පළ වී ඇත (Cavalli-Sforza et al. 1988; Soares et al. 2012a). එතැන් සිට නූතන මානවයන් අග්නිදිග ආසියාවේ සහ ඕෂනියාවේ ප්රදේශවලට වේගයෙන් ව්යාප්ත වී ඇතැයි සැලකේ (Forster & Matsumura 2005; Reed & Tishkoff 2006). නිදසුනක් වශයෙන්, ජාන සංචිත L3 හි පුද්ගලයන් අප්රිකාවේ සෝමාලි අර්ධද්වීපය හරහා මහාද්වීපයෙන් පිටතට සංක්රමණය වූ බව අධ්යයන දෙකකින් නිගමනය කර ඇත (Torroni et al. 2006; Soares et al. 2012b). එමෙන් ම මෙම ආකෘතිය යටතේ, එක් මානව කණ්ඩායමක් රතු මුහුද තරණය කර අරාබි අර්ධද්වීපයේ දකුණු වෙරළ තීරය දිගේ ඉන්දියාව දෙසට ගමන් කළ බව සැලකේ (Oppenheimer 2012a). මෙම කාල පරිච්ඡේදය තුළ සිදුවන ග්ලැසියර තත්ත්වයත් සමග මෙය අනුරූප වන අතර, එය රතු මුහුද හරහා ගමන් කිරීමට ඉඩ සලසන මුහුදු මට්ටම් පහත වැටීමට හේතු විය (Liu et al. 2006).
පුරාවිද්යාත්මක දෘෂ්ටිකෝණයකින් සලකා බැලූ විට, සමුද්රීය සම්පත් ගවේෂණය (Marine Exploitation) පෙන්නුම් කරන සාක්ෂි අතිශයින් සීමිතය. රතු මුහුදේ Abdur ගල්පරවලින් සහ ගල්ෆ් ද්රෝණියේ පුරාවිද්යාත්මක ස්ථානවලින් පුරාකෘති සොයා ගැනීම, එම ප්රදේශයේ අදින් වසර 100,000 පෙර දිගුකාලීන මානව ජනාවාස පෙන්නුම් කරයි; කෙසේ වෙතත්, මේවා නූතන මානව කණ්ඩායම්වල මුතුන් මිත්තන්ගේ ක්රියාකාරකම් නියෝජනය කරන්නේද යන්න තවමත් විවෘත ප්රශ්නයකි (Walter et al. 2000).
මානවයා අප්රිකාවෙන් පිට වී ගියේ කුමන කාලයක දීද?
තවත් ප්රධාන විවාදයක් වන්නේ අප්රිකාවෙන් පළමු මිනිසුන් පිටවීමේ නිශ්චිත කාලයයි. දැනට පිළිගත් මතයට අනුව යුරේසියාවට ලගා වීම අදින් වසර 50,000 – 60,000 අතර කාලයේ ද, නිශ්චිත වශයෙන්ම ඕස්ට්රේලියාවට ලගා වීම අදින් වසර 45,000- 50,000 ත් අතර කාලයේ සිදු විය (Stringer 2000; Mellars 2006; Underhill & Kivisild 2007; Shi et al. 2010; O’Connell & Allen 2012; Mellars et al. 2013). දෙවැන්න වන්නේ ටෝබා කන්ද (උතුරු සුමාත්රාවේ පිහිටි) පුපුරා යාමට පෙර අදින් වසර 74,000 දක්වා හා 100,000 – 130,000 පමණ වන විට මිනිසුන් අප්රිකාවෙන් පිටවී ගිය බව ය (Rampino & Self 1992; Mellars et al. 2013). මානවයා අප්රිකාවෙන් පිටවී යාම කාලනිර්ණය කිරීමේ දී ටෝබා පිපිරීම වැදගත් සන්දර්භයක් සපයයි, එනම්, නූතන සේපියන්ස් මානවයින් හා සම්බන්ධ ශිලාමෙවලම් එහි ගිනිකදු අළු තුළ සුරක්ෂිත වී ඇති බැවිනි (Petraglia et al. 2007). එනම් වසර 74,000කට පෙර නූතන මිනිසුන් අග්නිදිග ආසියාවට ළඟා වී ඇති බවට මෙය පැහැදිලි සාක්ෂි සපයයි. පුරාවිද්යා ස්ථානය වන Jebel Faya (වර්තමාන එක්සත් අරාබි එමීර් රාජ්යය) සහ Dhofar (වර්තමාන ඕමානය) හි නූබියන් සංකීර්ණයෙන් සොයාගත් ශිලා මෙවලම් අරාබිය හරහා සිදු වූ මුල් සංක්රමණය සඳහා තවදුරටත් සහාය ලබා දී ඇත (Armitage et al. 2011). එමෙන් ම, Daoxian (දකුණු චීනය) හි Fuyan ගුහාවෙන් අදින් වසර 80,000- 120,000 පැරණි මිනිස් දත් 47 ක් මෑතදී සොයා ගැනීම (Liu et al. 2015) තුළින්, ප්ලයිස්ටොසීන යුගයේ මුල් භාගයේදී නූතන මානවයා ආසියාව පුරා ව්යාප්ත වීම පිළිබද න්යායට සහාය වේ. කෙසේ වෙතත්, මෙම කණ්ඩායම් අතර සිටින පුද්ගලයින් නවීන ජනගහනයට ජානමය වශයෙන් දායක වූයේද නැතහොත් ලෙවන්ට් හි යෝජනා කර ඇති ආකාරයටම තවත් අසාර්ථක නික්මයාමක් නියෝජනය කරන්නේද යන්න සොයා බැලිය යුතුය. අදින් වසර 70,000- 120,000 ක කාලයට වඩා ඈත පෙරදිග නූතන මානවයා ස්ථානගත කිරීමට උපකාර විය හැකි අනෙකුත් පුරාවිද්යාත්මක සාක්ෂි වන්නේ Zhirendong (දකුණු චීනය) හි මානව අවශේෂ (Liu et al. 2010), ලුනා ගුහාවේ මෑත ප්ලයිස්ටොසීනයට අදාල මානව දත් (Bae et al. 2014), දකුණු චීනයේ Liujiang සැකිල්ල (Rosenberg 2002), හා Callao man නම් වූ අදින් වසර 67,000කට කාලනිර්ණය කර ඇති පිලිපීනයේ වූ මානව පාද අස්ථියයි (Mijares et al. 2010).
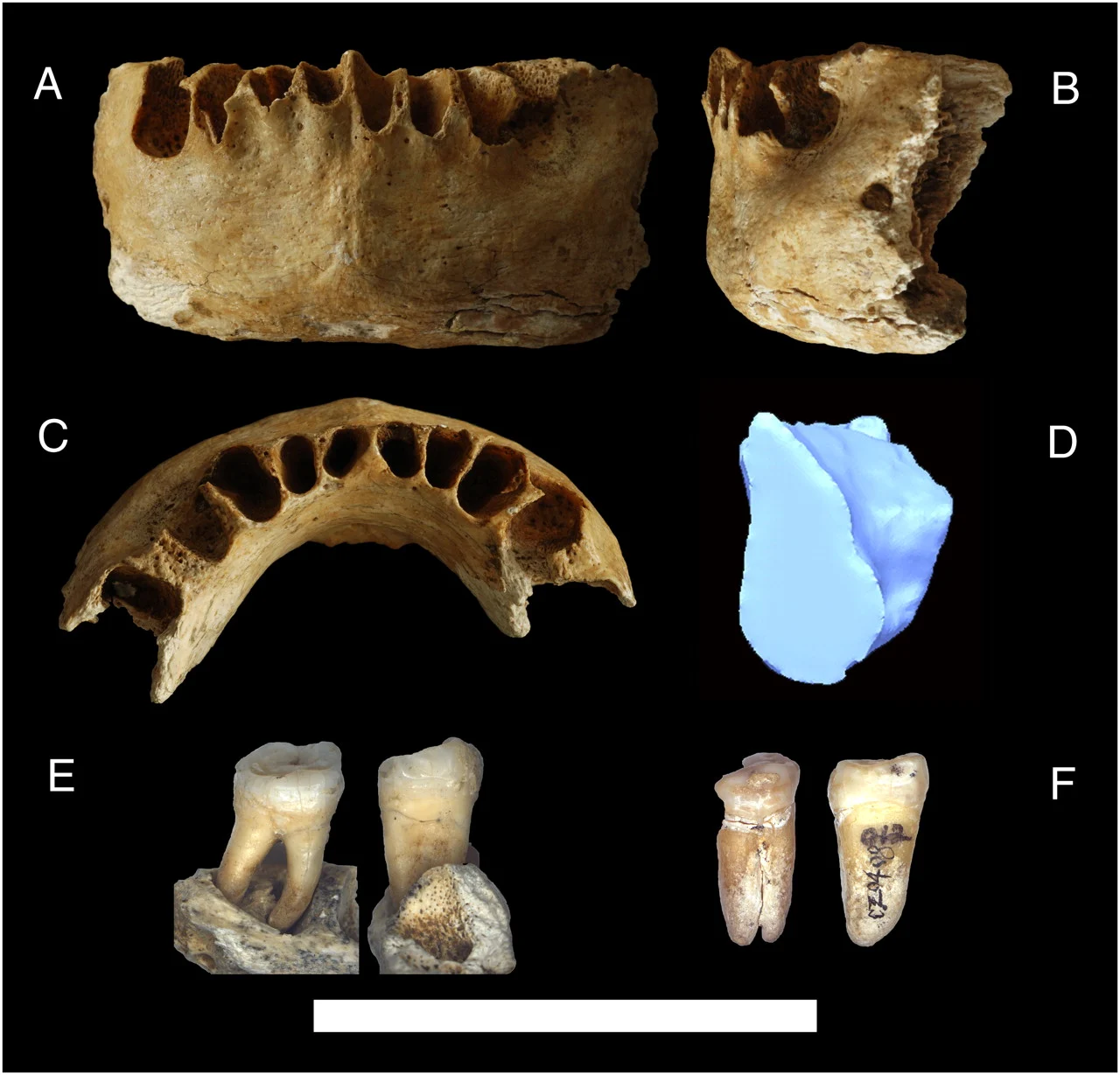
මෙම විවිධ දින සඳහා පරස්පර පුරාවිද්යාත්මක සාක්ෂි නිරාකරණය කිරීමට ජාන අධ්යයනයට තවමත් නොහැකි වී ඇති අතර, ඒවා පූර්ව ටෝබා (අදින් වසර 100,000-130,000) හෝ පශ්චාත් ටෝබා (අදින් වසර 50,000-60,000 පමණ) ලෙසද වර්ග කළ හැක. බොහෝවිට පෞරාණික ජාන (aDNA) දත්ත අවකාශීයව නොතිබීම හෝ සංරක්ෂණය නොවීම නිසා, එවැනි ජාන පදනම් වූ ප්රවේශ, නවීන ජාන භාවිතා කරමින් අනුමාන කිරීම කෙරෙහි විද්යාඥයින් අවධානය යොමු කර ඇත. මාතෘජාන සංචිත ප්රතිනිර්මාණය කිරීම මත පදනම් වූ අධ්යයනයන් අදින් වසර 60,000 සහ 40,000 අතර දිනයකදී නූතන මිනිසුන් අප්රිකාවෙන් පිට වූ බවට යෝජනා කර ඇත (Underhill & Kivisild 2007).
නියැන්ඩතාල් මිනිසුන් හා ඩෙනිසෝවන්වරුන් සමග මිශ්රවීම
ඔවුන් අප්රිකාවෙන් පිටතට සංක්රමණය වීමේදී, පිටත සේපියන්වරු පමණක් තනිවම නොසිටි අතර, අවම වශයෙන් යුරේසියාවේ වාසය කරන මානව කණ්ඩායම් දෙකක් එනම්, නියැන්ඩර්තාල් සහ ඩෙනිසෝවන් ලෙස පිටත ලෝකයේ විසී ය. ප්රතිස්ථාපන න්යාය හා අප්රිකාවෙන් පිට වී යාමේ න්යාය සාම්ප්රදායිකව සලකා බැලීමේ දී ජාන මිශ්රවීමක් පිළිබද අදහස් දැක්වීම අපහසු ය. නමුත් සංශෝධනය කරන ලද ප්රතිස්ථාපිත න්යාය අනුව අප්රිකාවෙන් පිටවී යාමේ දි සිදු වූ ජාන ගලනයක් සදහා උපකල්පන ගොඩනැගිය හැකි වේ, තවදුරටත් එය බහුප්රාදේශීය න්යායෙන් ව්යුක්තව දැක්විය නොහැකි වන අතර ම candelabra ආකෘතියෙන් වෙන් වී පවතී.
මානව පරිණාමයේ කතාන්දරයේ දී පුරාණ ජාන සමග සිදුකරන ලද පර්යේෂණවල වඩාත් උද්යෝගිමත් ප්රතිඵල වන්නේ නියැන්ඩතාල් මිනිසුන් හා ඩෙනිසෝවන් මානවයා හා සම්බන්ධ පර්යේෂණ පත්රිකා ය (Reich et al. 2010; Meyer et al. 2012; Prüfer et al. 2014). නියැන්ඩර්තාල් විශේෂය සොයාගත් ජර්මනියේ නියැන්ඩර් නිම්නයේ නමින් නම් කරන ලද නියැන්ඩර්තාල්, ප්රථම වරට අදින් වසර ලක්ෂ 2 සිට 2.5 දක්වා කාලයට අයත් ලෙස සැලකේ (Hublin 2009), නමුත් නිශ්චිත පරාසය තවමත් විවාදයට ලක්ව ඇති අතර, අවම වශයෙන් දකුණු යුරෝපයේ ප්රදේශ වල, ඔවුන් අදින් වසර 30,000 දක්වාම වන තෙක් පැවතුනි (Finlayson et al. 2006). මූලික ජාන විශ්ලේෂණ මාතෘජාන සංචිත කෙරෙහි අවධානය යොමු කරන ලද අතර, එය පැරණි සාම්පල වලින් වඩාත් පහසුවෙන් නිස්සාරණය කර විස්තාරණය කළ හැකි අතර (extracted and amplified), යුරේසියාවට සංක්රමණය වන විට නියැන්ඩර්තාල් සහ නූතන මානවයන් අතර අන්තර් මිශ්ර නොවීයැයි යෝජනා කරන ලදී (Krings et al. 1997a; b; Serre et al. 2004; Green et al. 2008).

දකුණු සයිබීරියාවේ Altai කඳුකරයේ ඩෙනිසෝවා ගුහාවෙන් සොයා ගන්නා ලද මානව ඇගිලි පුරුක විශ්ලේෂණයෙන් ඩෙනිසෝවන් නම් වූ නියැන්ඩර්තාල් මානවයන් හා සම්බන්ධ ජානමය වශයෙන් වෙනස් වූ පුරාවිද්යා මානව සමූහයක පැවැත්ම තහවුරු විය (Reich et al. 2010; Meyer et al. 2012). ඩෙනිසෝවන් පරම්පරාව ප්රවේණික සාක්ෂි මත පදනම්ව වර්ගීකරණය කරන ලද අතර නියැන්ඩර්තාල්වරුන් වෙතින් ජානමය වශයෙන් අපසරණය වී ඇති බව ඇස්තමේන්තු කර ඇත (Prüfer et al. 2014). එහි එන විශ්මයජනක සොයාගැනීමක් නම්, සයිබීරියාවේදී සොයා ගෙන ඇති, ඩෙනිසෝවන් ජාන නවගිනියානුවන්, ඕස්ට්රේලියානුවන් සහ ඇතුළු නවීන දින ආසන්න සාගර වැසියන් සමඟ පොදුවේ හඳුනාගත හැකි ජාන කොටස් (3%-5% ක ප්රමාණයක්) බෙදා ගන්නා බව සොයා ගන්නා ලදී (Finlayson et al. 2006; Reich et al. 2010, 2011; Meyer et al. 2012). එතැන් සිට, ඩෙනිසෝවන් ජාන නූතන ජනගහනය තුළ තවදුරටත් අනාවරණය වී ඇත, උදාහරණයක් ලෙස, නැගෙනහිර යුරේසියාවේ සහ ස්වදේශික ඇමරිකාවේ නූතන ජනගහනයේ ඩෙනිසෝවන් ජාන පුළුල් ලෙස පැතිරී ඇති බව යෝජනා කරයි (Prüfer et al. 2014; Qin & Stoneking 2015). නිදසුනක් වශයෙන්, ඉහළ උන්නතාංශවල වෙසෙන ටිබෙට් වැසියන්ගේ ඔක්සිජන් රහිත තත්වයන්ට අනුවර්තනය වන ජාන සංචිත වර්ග ඩෙනිසෝවන් හෝ ඩෙනිසෝවන් ආශ්රිත පුද්ගලයින්ගේ ජාන ව්යුහයේ ද දක්නට ඇති අතර, එය අනෙකුත් මානව වර්ග සමඟ මානව ජාන විවිධත්වය රැකගැනීමේ දී අනුවර්තනය වීමේ දී වැදගත් විය හැකි බව පර්යේෂකයින් යෝජනා කරයි (Huerta-Sánchez et al. 2014).
සංක්රමණ තරංග: තනි හෝ බහුවිධ අවස්ථා?
මානවයා අප්රිකාවෙන් පිටවී යාමේ න්යාය හා සම්බන්ධව පවතින මතභේදයේ තවත් මාතෘකාවක් වන්නේ අප්රිකාවෙන් පිටව ගිය ප්රථම නූතන මානවයන් එක් වරකදී පමණක් පිටවී ගියා ද නැතහොත් ඇත්ත වශයෙන්ම ලෝකයේ සෙසු ප්රදේශවල ජනාකීර්ණ කිරීම සඳහා අප්රිකාවෙන් පිටතට විසිරී යාමේ වාර කිහිපයක් තිබේද යන්නයි. ඒ සදහා විවිධ විසරණ ආකෘති පවතින අතර, ඒ සියල්ලම විද්යාත්මකව තහවුරු කරන ලද ඒවා ය (Groucutt et al. 2015b).
මානවයා අප්රිකාවෙන් පිට වී, යුරේසියාවේ සෙසු ප්රදේශවලට තනි තරංගයක් ලෙස උතුරේ හෝ දකුණේ හෝ තනි මාර්ගයක් හරහා සංක්රමණය වූ බව තනි විසුරුම් (single dispersal) ආකෘතියෙන් දැක්වේ. නමුත් තනි විසුරුමේ ආකෘතියේ ප්රභේද ඒවායේ කාලසීමාව අනුව වෙනස් වේ, නිදසුනක් ලෙස, ඉහළ පැලියොලිතික් ආකෘතිය යටතේ සම්පූර්ණ ජනගහන ප්රතිස්ථාපනය සමඟින් අදින් වසර 50,000ට පසුවත් (Shea 2001; Meignen 2012; Kuhn 2013), Skhul සහ Qafzeh මානව අවශේෂ සහ දේශගුණික දත්ත මත පදනම්ව මානවයාගේ පිටවී යාම අදින් වසර 100,00-120,000 දක්වාත් උපකල්පනය කෙරේ (McDermott et al. 1993; Frumkin et al. 2011). තවදුරටත්, අදින් වසර 50,000- 75,000කට පෙර දිනයකදී (Oppenheimer 2012b) මානවයින් ගමන්ගත් මගේ සන්දර්භය අනුව ද විවාදය වෙනස් මගත් ගනී, එනම් ඔවුන් වෙරළබඩ කාලපය දිගේ ගමන් ගත් බවට වර්තමාන එක්සත් අරාබි අරාබියේ හා ඊසානදිග අප්රිකාවේ ඇති පුරාකෘති මත පදනම්ව අදහස් පළ වී ඇත (Armitage et al. 2011a; Usik et al. 2013).
මෙම සියලු ආකෘතීන් යටතේ, ජානමය සාක්ෂිවලින් පෙනී යන්නේ අප්රිකාවෙන් පිටතට සංක්රමණය වීම ආරම්භක සංක්රමණ කණ්ඩායම් තුළ දැඩි බාධකයක් සමඟ ඇති වූ බවත්, වේගවත් ප්රසාරණයකට පෙර ජාන විවිධත්වය විශාල ලෙස අඩු කරන බවත්ය. ඒ අනුව අනුක්රමික නිර්මාතෘ ප්රයෝග ආකෘතිය (serial founder effect model) යනු තනි විසුරුමේ ආකෘතියක් පිළිබඳ විය හැකි එක් අර්ථකථනයක් වන අතර (Deshpande et al. 2009), අප්රිකාවෙන් පිටතට විසිරීම මුලින් සිදු වූයේ තනි සංක්රමණික කණ්ඩායමක් හරහා වන අතර පුද්ගලයන් ජනාකීර්ණ ප්රදේශවලට විසිරී, ජනගහන ප්රමාණයෙන් ව්යාප්ත වූ අතර, ඉහළ නැංවීමක් ලෙස පුනරාවර්තන ආකාරයෙන් දිගටම පැවතුනි. මෙම අනුක්රමික නිර්මාතෘ ක්රියාවලිය අප්රිකාවේ සිට භූගෝලීය දුර වැඩි වන විට කණ්ඩායම් වශයෙන් නිරීක්ෂණය කරන ලද ජානමය ප්ලාවිතය වැඩි වීමේ සහ ජාන විවිධත්වය අඩු වීමේ රටාවන් ඇති කළ හැකිය (Pemberton et al. 2013).
නවතම සොයාගැනීම් අනුව ජානවිවිධත්වය සහ භූගෝලීය දුර අතර නිරීක්ෂණය කරන ලද ප්රතිලෝම සහසම්බන්ධය තවදුරටත් ඓතිහාසික ආකෘති යටතේ උත්පාදනය කළ හැකි බව පෙන්වා දී ඇත (Pickrell & Reich 2014). පළමුවෙන්ම නූතන මානවයා අප්රිකාවෙන් පිටවී යාමේ ස්වභාවය සලකා බලන විට නැගෙනහිර ආසියාව ප්රධාන කලාපයකි. තනි විසුරුමේ ආකෘතියක් යටතේ, නැගෙනහිර ආසියාවේ ජනතාව තනි රැල්ලක් හරහා සිදු වූ බවට උපකල්පනය කර ඇත, උදාහරණයක් ලෙස, ආදිවාසී ඕස්ට්රේලියානුවන් මෙම නැඟෙනහිර ආසියානු පුද්ගලයන්ගේ රැල්ල තුළින් විවිධාංගීකරණය වී ඇතැයි අනුමාන කෙරේ. මෑත කාලීන සම්පූර්ණ-ප්රවේණික අධ්යයන හෙළි කරන්නේ යුරෝපීයයන් සහ ආසියානුවන් අතර අදින් වසර 17,000- 43,000 අතර කාලයකදී සිදුවූ ජානමය බෙදීමක් පිළිබදව ය (Keinan et al. 2009; Gutenkunst et al. 2009). නමුත් එම සොයාගැනීම් ඕස්ට්රේලියාව, නිව්ගිනියාව හා මෙලනීසියාව හි හමුවන පුරාවිද්යාත්මක සාක්ෂි සමග ඝට්ටනය වන බව පෙනේ, එනම් අදින් වසර 50,000කට පෙර මෙම කලාපයේ මානවයා ජනාවාස ගතව සිටි බව පුරාවිද්යාත්මක සාක්ෂි මගින් සනාථ වේ (Bowler et al. 1970).
එක් ප්රසාරණ තරංගයකින් නොව බහුවිධ වාර හරහා කායච්ඡේදවිද්යාත්මකව නූතන මානවයා ලෝකය ජනාවාස කළ බව මුලින්ම යෝජනා කළේ වර්ෂ 1994 දී Lahr සහ Foley ය (Lahr & Foley 1998, 2005). ඔවුන්ගේ බහු විසුරුම් ආකෘතිය යටතේ, දකුණු අරාබිය හරහා අග්නිදිග ආසියාවට (දකුණු මාර්ගය) අදින් වසර 50,000 -100,000 කාලයේ දී මුල් විසරණයක් සිදු වූ අතර, අදින් වසර 40,000- 50,000 කාලයේ දී උතුරු මාර්ගය ඔස්සේ ලෙවන්ට් හරහා දෙවන සංක්රමණය යුරේසියාවේ සෙසු ප්රදේශ ජනාවාසකරණයට හේතු විය. තවදුරටත් මධ්ය පුරාශිලායුගයට අයත් ගල් ආයුධ සමඟ අනුබද්ධ අරාබි අර්ධද්වීපයට පළමු සංක්රමණය සිදුවීම සහ පසුකාලීනව Levantine කොරිඩෝව හරහා යුරෝපයට ව්යාප්ත වීම ඉහළ පැලියෝලිතික් මෙවලම්වල පෙනුමට සමග සමකාලීන බව බැවින්, එම එක් එක් විසිරීම් වාර ඒ හා සම්බන්ධ පුරාකෘති අනුව වෙනස් වන බව ඔවුන් යෝජනා කළහ. පසුකාලීනව Field සහ Lahr විසින් සිදු කරන ලද පර්යේෂණ තුළින්, භූගෝලීය තොරතුරු පද්ධතිය පදනම් කරගත් ආකෘතියක් භාවිතා කරමින්, පාරිසරික බාධක මත අප්රිකාවෙන් උතුරු සහ දකුණු මාර්ග යන ද්විත්වයම භාවිත කළ හැකි බව පෙන්වීමට අදින් වසර 74,000- 159,000 දක්වා වූ දේශගුණික දත්ත උපයෝගී කරගන්නා ලදී (Field & Lahr 2005).
අග්නිදිග ආසියාවේ වූ නූතන මානවයාගේ ජනාවාසකරණය බෙහෙවින් විවාදාත්මක මාතෘකාවකි. එහි දී අවසන් ග්ලැසියර උපරිමය හා නවශිලායුගයට අයත් මුහුදු මට්ටම් විචලනය වීම අතිශය තීරණාත්මක සාධක දෙකකි. 2017 වර්ෂයේ දී කායච්ඡේද විද්යාත්මකව නූතන මානවයාගේ දන්තීන හැඩය හා චිචල්යතාවය පරීක්ෂා කර, එම ප්රදේශයේ නියෝලිතික ගොවීන්ගේ ජීවවිද්යාත්මක පැවැත්ම සදහා එක් ජාන සමූහයකින් නොව එකිනෙකට වෙනස් ජානවර්ග කීපයකම ආභාෂය ලැබී ඇති බව නිගමනය වී තිබේ (Corny et al. 2017).
සමාලෝචනය
පළමු හෝමෝ සේපියන්ස් මානවයාගේ මූලාරම්භය සහ සංක්රමණික චලනයන් හෙළිදරව් කිරීම නූතන ජනගහනයේ ජාන විවිධත්වය අවබෝධ කර ගැනීම සඳහා ප්රධාන වේ. එනම්, කාලයාගේ ඇවෑමෙන්, පුරාපාෂාණ විද්යාත්මක, පුරාවිද්යාත්මක සහ දේශගුණික දත්ත සංශ්ලේෂණය කිරීමේදී බොහෝ විට ගැටුම් ඇති වන සංකීර්ණතා මට්ටමක් ජනනය කරයි. අවසාන දශකය තුළ සිදුවී ඇති පර්යේෂණ සැලකීමේ දී ඒවා බොහෝවිට සාධක තනිව ගෙන විශ්ලේෂණය කිරීම වෙනුවය අන්තර්විෂයාත්මක හා බහුවිෂයාත්මක ප්රවේශයක් භාවිත කිරීම නිරීක්ෂණය කළ හැකි ය. එනම්, අතීත සිදුවීම් ප්රතිනිර්මාණය කිරීම සඳහා ජානිවද්යාත්මක දත්ත භාවිතා කිරීම කෙරෙහි අවධානය යොමු කරයි. භූගෝලීය වශයෙන් සහ තාවකාලිකව නියෝජනය වන කලාපවලින් ඉහළ ආවරණයක් හෝ පැතිරීමක් සහිත aDNA සාම්පල නොමැතිකම හේතුවෙන්, බොහෝ අධ්යයනයන් අප්රිකාවෙන් පිටවීමේ න්යායට අදාළ ප්රශ්නවලට පිළිතුරු සැපයීම සඳහා නවීන ජනගහන ආශ්රිත දත්ත භාවිතා කිරීම කෙරෙහි අවධානය යොමු කර ඇත. නූතන ජනගහනයෙන් බහුතරයක් මිශ්ර වී ඇති අතර බොහෝ අප්රිකානු ජනගහණයන්ට අප්රිකානු නොවන සම්භවයක් ලැබී ඇත්තේ නැවත අප්රිකාවට සංක්රමණය වීම නිසා මෙය අතිශයින් අභියෝගාත්මක ය. තවද, ජනගහන-විශේෂිත ප්ලාවිත බලපෑම් (population-specific drift effects) වැදගත් වනු ඇත, විශේෂයෙන්ම අප්රිකාවෙන් පිටතට තනි හෝ බහු විසුරුවීම් පිළිබඳ ප්රශ්නය ආමන්ත්රණය කිරීමේදී, විශේෂයෙන් යුරෝපීයයන්ට සාපේක්ෂව නැගෙනහිර ආසියාතිකයන්ගේ ඉහළ ප්ලාවිතය ලබා දී ඇති විට, අප්රිකාවට සාපේක්ෂව බෙදීම් කාලය පිළිබඳ ඇස්තමේන්තු විකෘති විය හැකිය.
අපි දැන් පිවිසෙමින් සිටින්නේ විවිධ ප්රදේශවලින් සහ කාල ස්ථානවලින් aDNA සාම්පල වැඩි වැඩියෙන් ලබා ගත හැකි යුගයකට ය. aDNA ප්රතිසාධනයේ නව ශිල්පීය ක්රම දියුණු කිරීම සහ දූෂණය අවම කිරීම සඳහා ප්රවේශයන් වැඩිදියුණු කිරීම සහ පශ්චාත් පරීක්ෂණ හානිය සඳහා පර්යේෂකයන්ට දැන් ප්රමාණවත් තත්ත්වයේ ජාන වෙත ප්රවේශ වීමට සහ වඩාත් සංකීර්ණ විශ්ලේෂණ ක්රමවේද සඳහා ප්රවේශය ඇත. මේ අනුව, මානවයා අප්රිකාවෙන් පිටවී ගියේ කවදාද, කුමන භූගෝලීය ප්රදේශයක් හරහා ද, එක වරකදී හෝ වාර කීපයකදී පිටවී ගියේ ද යන ප්රශ්නවලට අදාළ තොරතුරු දත්ත ලබා ගත හැකිද යන්න සොයා බැලිය යුතුය. මෙම දිගුකාලීන වාද විවාදවලින් සමහරක් විසඳීමේ අනාගත සාර්ථකත්වය සඳහා පුරාවිද්යාත්මක හා මානව විද්යාත්මක දත්තවල වර්ධනය වෙමින් පවතින න්යායාත්මක අගයන් මෙන්ම, පෞරාණික හා නවීන ජානවල නිරීක්ෂිත රටා ඒකාබද්ධ කරන බහුවිෂයාත්මක ප්රවේශයක් අවශ්ය වනු ඇත.
කෙටි වාර්තාමය චිත්රපට
- Human Origins: Out of Africa Theory Debunked?
- Children of Adam | National Geographic DNA Documentary
- When And Why Did Our Early Ancestors Migrate Out Of Africa
- Is the Out of Africa Model Wrong?
- Debunking Out-of-Africa Theory in Under 15 Minutes
- “Out of Africa” Theory Changed
මනුෂ්යත්වයේ කතාන්දරය: මානව පරිණාමය
- මානව පරිණාමයේ ගමන 1: මනුෂ්යත්වයේ ආරම්භය
- මානව පරිණාමයේ ගමන 2- මනුෂ්යත්වයේ මූල බීජ: මුල් වානර මානවයින් හා ඔස්ට්රලෝපිතිකස් ගණය
- මානව පරිණාමයේ ගමන 3- මුල් හෝමෝ සාමාජිකයින්: හෝමෝ හැබිලිස් හා හෝමෝ ඉරෙක්ටස්
- මානව පරිණාමයේ ගමන 4: හෝමෝ නියැන්ඩතාල් මානවයා
- මානව පරිණාමයේ ගමන 5: මානව නූතනත්වය: හෝමෝ සේපියන් මානවයාගේ සම්භවය
- මානව පරිණාමයේ ගමන 6: මානව විවිධත්වය හා නව මානව විශේෂ
- මානව පරිණාමයේ නොනවතින සංවාදය 7: නූතන මානවයා අප්රිකාවෙන් පිට වී යාම
- මානවයා නවලොව ජනාවාසකරණය කිරීම (අමරිකාව හා ඕස්ට්රේලියාව)
- නියෝලිතික විප්ලවය හා මුල් කෘෂිකර්මයේ නැගීම
References
Alonso, S. & J.A.L. Armour. 2001. A highly variable segment of human subterminal 16p reveals a history of population growth for modern humans outside Africa. Proceedings of the National Academy of Sciences 98: 864–69.
Armitage, S.J., S.A. Jasim, A.E. Marks, A.G. Parker, V.I. Usik & H.-P. Uerpmann. 2011a. The Southern Route “Out of Africa”: Evidence for an Early Expansion of Modern Humans into Arabia. Science 331: 453–56.
Bae, C.J., W. Wang, J. Zhao, S. Huang, F. Tian & G. Shen. 2014. Modern human teeth from Late Pleistocene Luna Cave (Guangxi, China). Quaternary International 354: 169–83.
Bowler, J.M., R. Jones, H. Allen & A.G. Thorne. 1970. Pleistocene human remains from Australia: A living site and human cremation from Lake Mungo, western New South Wales. World Archaeology 2: 39–60.
Cann, R.L., M. Stoneking & A.C. Wilson. 1987. Mitochondrial DNA and human evolution. Nature 325: 31–36.
Caramelli, D. et al. 2003. Evidence for a genetic discontinuity between Neandertals and 24,000-year-old anatomically modern Europeans. Proceedings of the National Academy of Sciences 100: 6593–97.
Cavalli-Sforza, L.L. & M.W. Feldman. 2003. The application of molecular genetic approaches to the study of human evolution. Nature Genetics 33: 266–75.
Cavalli-Sforza, L.L., A. Piazza, P. Menozzi & J. Mountain. 1988. Reconstruction of human evolution: bringing together genetic, archaeological, and linguistic data. Proceedings of the National Academy of Sciences 85: 6002–6.
Clark, J.D. et al. 2003. Stratigraphic, chronological and behavioural contexts of Pleistocene Homo sapiens from Middle Awash, Ethiopia. Nature 423: 747–52.
Coon, C.S. 1962. The Origin of Races. New York: Knopf.
Corny, J. et al. 2017. Dental phenotypic shape variation supports a multiple dispersal model for anatomically modern humans in Southeast Asia. Journal of Human Evolution 112: 41–56.
Darwin C. 1871. The Descent of Man, and Selection in Relation to Sex. London: John Murray.
Deshpande, O., S. Batzoglou, M.W. Feldman & L. Luca Cavalli-Sforza. 2009. A serial founder effect model for human settlement out of Africa. Proceedings of the Royal Society B: Biological Sciences 276: 291–300.
Fadhlaoui-Zid, K., M. Haber, B. Martínez-Cruz, P. Zalloua, A. Benammar Elgaaied & D. Comas. 2013. Genome-Wide and Paternal Diversity Reveal a Recent Origin of Human Populations in North Africa. PLoS ONE 8: 25-26.
Field, J.S. & M.M. Lahr. 2005. Assessment of the Southern Dispersal: GIS-Based Analyses of Potential Routes at Oxygen Isotopic Stage 4. Journal of World Prehistory 19: 1–45.
Finlayson, C. et al. 2006. Late survival of Neanderthals at the southernmost extreme of Europe. Nature 443: 850–53.
Forster, P. & S. Matsumura. 2005. Did Early Humans Go North or South? Science 308: 965–66.
Frumkin, A., O. Bar-Yosef & H.P. Schwarcz. 2011. Possible paleohydrologic and paleoclimatic effects on hominin migration and occupation of the Levantine Middle Paleolithic☆. Journal of Human Evolution 60: 437–51.
Green, R.E. et al. 2008. A Complete Neandertal Mitochondrial Genome Sequence Determined by High-Throughput Sequencing. Cell 134: 416–26.
Groucutt, H.S. et al. 2015a. Human occupation of the Arabian Empty Quarter during MIS 5: evidence from Mundafan Al-Buhayrah, Saudi Arabia. Quaternary Science Reviews 119: 116–35.
—. 2015b. Rethinking the dispersal of Homo sapiens out of Africa. Evolutionary Anthropology: Issues, News, and Reviews 24: 149–64.
Grün, R. et al. 2005. U-series and ESR analyses of bones and teeth relating to the human burials from Skhul. Journal of Human Evolution 49: 316–34.
Gutenkunst, R.N., R.D. Hernandez, S.H. Williamson & C.D. Bustamante. 2009. Inferring the Joint Demographic History of Multiple Populations from Multidimensional SNP Frequency Data. PLoS Genetics 5: 13-18.
Hawks, J. 2001. The Y Chromosome and the Replacement Hypothesis. Science 293: 567–567.
Henn, B.M. et al. 2011. Hunter-gatherer genomic diversity suggests a southern African origin for modern humans. Proceedings of the National Academy of Sciences 108: 5154–62.
Horai, S., K. Hayasaka, R. Kondo, K. Tsugane & N. Takahata. 1995. Recent African origin of modern humans revealed by complete sequences of hominoid mitochondrial DNAs. Proceedings of the National Academy of Sciences 92: 532–36.
Hublin, J.J. 2009. The origin of Neandertals. Proceedings of the National Academy of Sciences 106: 16022–27.
Huerta-Sánchez, E. et al. 2014. Altitude adaptation in Tibetans caused by introgression of Denisovan-like DNA. Nature 512: 194–97.
Huxley, T. 1863. Evidence as to Man’s Place in Nature. London: Williams & Norgate.
Ingman, M., H. Kaessmann, S. Pääbo & U. Gyllensten. 2000. Mitochondrial genome variation and the origin of modern humans. Nature 408: 708–13.
Keinan, A., J.C. Mullikin, N. Patterson & D. Reich. 2009. Accelerated genetic drift on chromosome X during the human dispersal out of Africa. Nature Genetics 41: 66–70.
Kim, H.L., A. Ratan, G.H. Perry, A. Montenegro, W. Miller & S.C. Schuster. 2014. Khoisan hunter-gatherers have been the largest population throughout most of modern-human demographic history. Nature Communications 5: 5692.
Krings, M., A. Stone, R.W. Schmitz, H. Krainitzki, M. Stoneking & S. Pääbo. 1997a. Neandertal DNA Sequences and the Origin of Modern Humans. Cell 90: 19–30.
Kuhn, S.L. 2013. Roots of the Middle Paleolithic in Eurasia. Current Anthropology 54: S255–68.
Lahr, M.M. & R. Foley. 2005. Multiple dispersals and modern human origins. Evolutionary Anthropology: Issues, News, and Reviews 3: 48–60.
Lahr, M.M. & R.A. Foley. 1998. Towards a theory of modern human origins: geography, demography, and diversity in recent human evolution. American journal of physical anthropology Suppl 27: 137–76.
Liu, H., F. Prugnolle, A. Manica & F. Balloux. 2006. A Geographically Explicit Genetic Model of Worldwide Human-Settlement History. The American Journal of Human Genetics 79: 230–37.
Liu, W. et al. 2010. Human remains from Zhirendong, South China, and modern human emergence in East Asia. Proceedings of the National Academy of Sciences 107: 19201–6.
—. 2015. The earliest unequivocally modern humans in southern China. Nature 526: 696–99.
Luis, J.R., D.J. Rowold, M. Regueiro, B. Caeiro, C. Cinnioğlu, C. Roseman, P.A. Underhill, L.L. Cavalli-Sforza & R.J. Herrera. 2004. The Levant versus the Horn of Africa: Evidence for Bidirectional Corridors of Human Migrations. The American Journal of Human Genetics 74: 532–44.
Macaulay, V. et al. 2005. Single, Rapid Coastal Settlement of Asia Revealed by Analysis of Complete Mitochondrial Genomes. Science 308: 1034–36.
McCarthy, R.C. & L. Lucas. 2014. A morphometric re-assessment of BOU-VP-16/1 from Herto, Ethiopia. Journal of Human Evolution 74: 114–17.
McDermott, F., R. Grün, C.B. Stringer & C.J. Hawkesworth. 1993. Mass-spectrometric U-series dates for Israeli Neanderthal/early modern hominid sites. Nature 363: 252–55.
McDougall, I., F.H. Brown & J.G. Fleagle. 2005. Stratigraphic placement and age of modern humans from Kibish, Ethiopia. Nature 433: 733–36.
Meignen, L. 2012. Levantine Perspectives on the Middle to Upper Paleolithic “Transition.” Archaeology, Ethnology and Anthropology of Eurasia 40: 12–21.
Mellars, P. 2006. Going East: New Genetic and Archaeological Perspectives on the Modern Human Colonization of Eurasia. Science 313: 796–800.
Mellars, P., K.C. Gori, M. Carr, P.A. Soares & M.B. Richards. 2013. Genetic and archaeological perspectives on the initial modern human colonization of southern Asia. Proceedings of the National Academy of Sciences 110: 10699–704.
Metspalu, M. et al. 2004. Most of the extant mtDNA boundaries in south and southwest Asia were likely shaped during the initial settlement of Eurasia by anatomically modern humans. BMC genetics 5: 26.
Meyer, M. et al. 2012. A High-Coverage Genome Sequence from an Archaic Denisovan Individual. Science 338: 222–26.
Mijares, A.S. et al. 2010. New evidence for a 67,000-year-old human presence at Callao Cave, Luzon, Philippines. Journal of Human Evolution 59: 123–32.
O’Connell, James.F. & J. Allen. 2012. Forum The Restaurant At The End Of The Universe: Modelling the colonisation of Sahul. Australian Archaeology 74: 5–31.
Oppenheimer, S. 2012a. Out-of-Africa, the peopling of continents and islands: tracing uniparental gene trees across the map. Philosophical Transactions of the Royal Society B: Biological Sciences 367: 770–84.
—. 2012b. A single southern exit of modern humans from Africa: Before or after Toba? Quaternary International 258: 88–99.
Ovchinnikov, I. v., A. Götherström, G.P. Romanova, V.M. Kharitonov, K. Lidén & W. Goodwin. 2000. Molecular analysis of Neanderthal DNA from the northern Caucasus. Nature 404: 490–93.
Pagani, L. et al. 2015. Tracing the Route of Modern Humans out of Africa by Using 225 Human Genome Sequences from Ethiopians and Egyptians. The American Journal of Human Genetics 96: 986–91.
Pemberton, T.J., M. DeGiorgio & N.A. Rosenberg. 2013. Population Structure in a Comprehensive Genomic Data Set on Human Microsatellite Variation. G3 Genes|Genomes|Genetics 3: 891–907.
Petraglia, M. et al. 2007. Middle Paleolithic Assemblages from the Indian Subcontinent Before and After the Toba Super-Eruption. Science 317: 114–16.
Pickrell, J.K. & D. Reich. 2014. Toward a new history and geography of human genes informed by ancient DNA. Trends in Genetics 30: 377–89.
Pope, K.O. & J.E. Terrell. 2007. Environmental setting of human migrations in the circum-Pacific region. Journal of Biogeography 12: 45-56.
Prüfer, K. et al. 2014. The complete genome sequence of a Neanderthal from the Altai Mountains. Nature 505: 43–49.
Prugnolle, F., A. Manica & F. Balloux. 2005. Geography predicts neutral genetic diversity of human populations. Current Biology 15: 159–60.
Qin, P. & M. Stoneking. 2015. Denisovan Ancestry in East Eurasian and Native American Populations. Molecular Biology and Evolution 32: 2665–74.
Ramachandran, S., O. Deshpande, C.C. Roseman, N.A. Rosenberg, M.W. Feldman & L.L. Cavalli-Sforza. 2005. Support from the relationship of genetic and geographic distance in human populations for a serial founder effect originating in Africa. Proceedings of the National Academy of Sciences 102: 15942–47.
Rampino, M.R. & S. Self. 1992. Volcanic winter and accelerated glaciation following the Toba super-eruption. Nature 359: 50–52.
Reed, F.A. & S.A. Tishkoff. 2006. African human diversity, origins and migrations. Current Opinion in Genetics & Development 16: 597–605.
Reich, D. et al. 2010. Genetic history of an archaic hominin group from Denisova Cave in Siberia. Nature 468: 1053–60.
—. 2011. Denisova Admixture and the First Modern Human Dispersals into Southeast Asia and Oceania. The American Journal of Human Genetics 89: 516–28.
Relethford, J.H. 2001. Absence of regional affinities of Neandertal DNA with living humans does not reject multiregional evolution. American Journal of Physical Anthropology 115: 95–98.
Relethford, J.H. 2008. Genetic evidence and the modern human origins debate. Heredity 100: 555–63.
Rito, T., M.B. Richards, V. Fernandes, F. Alshamali, V. Cerny, L. Pereira & P. Soares. 2013. The First Modern Human Dispersals across Africa. PLoS ONE 8: 67-96.
Rosenberg, Karen. 2002. A late Pleistocene human skeleton from Liujiang, China suggests regional population variation in sexual dimorphism in the human pelvis. Variability and Evolution 10: 5–17.
Schlebusch, C.M. et al. 2012. Genomic Variation in Seven Khoe-San Groups Reveals Adaptation and Complex African History. Science 338: 374–79.
Serre, D., A. Langaney, M. Chech, M. Teschler-Nicola, M. Paunovic, P. Mennecier, M. Hofreiter, G. Possnert & S. Pääbo. 2004. No Evidence of Neandertal mtDNA Contribution to Early Modern Humans. PLoS Biology 2: 57-86.
Shea, J.J. 2001. The archaeology of an illusion: the middle-upper paleolithic transition in the Levant, in J.M. le Tenesor, R. Jagher & R. Otte (ed.) The Lower and Middle Paleolithic in the Middle East and Neighbouring Regions.: 169–82. ERAUL 126. Liége: Université de Liége.
Shi, W. et al. 2010. A Worldwide Survey of Human Male Demographic History Based on Y-SNP and Y-STR Data from the HGDP-CEPH Populations. Molecular Biology and Evolution 27: 385–93.
Smith, T.M., P. Tafforeau, D.J. Reid, R. Grün, S. Eggins, M. Boutakiout & J.-J. Hublin. 2007. Earliest evidence of modern human life history in North African early Homo sapiens. Proceedings of the National Academy of Sciences 104: 6128–33.
Soares, P. et al. 2012a. The Expansion of mtDNA Haplogroup L3 within and out of Africa. Molecular Biology and Evolution 29: 915–27.
Stringer, C. 1992. Replacement, continuity and the origin of Homo sapiens, in G. Brauer & F.H. Smith (ed.) Continuity or Replacement? Controversies in Homo sapiens Evolution: 9–24. Rotterdam: Balkema.
Stringer, C. 2000. Coasting out of Africa. Nature 405: 25–27.
—. 2002. Modern human origins: progress and prospects. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences 357: 563–79.
Stringer, C.B. & P. Andrews. 1988. Genetic and fossil evidence for the origin of modern humans. Science (New York, N.Y.) 239: 1263–68.
Takahata, N., S.-H. Lee & Y. Satta. 2001. Testing Multiregionality of Modern Human Origins. Molecular Biology and Evolution 18: 172–83.
Tattersall, I. 2009. Human origins: Out of Africa. Proceedings of the National Academy of Sciences 106: 16018–21.
Templeton, A.R. 2007. Genetics and recent human evolution. Evolution; international journal of organic evolution 61: 1507–19.
Thomson, R., J.K. Pritchard, P. Shen, P.J. Oefner & M.W. Feldman. 2000. Recent common ancestry of human Y chromosomes: Evidence from DNA sequence data. Proceedings of the National Academy of Sciences 97: 7360–65.
Torroni, A., A. Achilli, V. Macaulay, M. Richards & H.-J. Bandelt. 2006. Harvesting the fruit of the human mtDNA tree. Trends in genetics : TIG 22: 339–45.
Underhill, P.A. & T. Kivisild. 2007. Use of Y Chromosome and Mitochondrial DNA Population Structure in Tracing Human Migrations. Annual Review of Genetics 41: 539–64.
Underhill, P.A. et al. 2000. Y chromosome sequence variation and the history of human populations. Nature Genetics 26: 358–61.
Usik, V.I., J.I. Rose, Y.H. Hilbert, P. van Peer & A.E. Marks. 2013. Nubian Complex reduction strategies in Dhofar, southern Oman. Quaternary International 300: 244–66.
Vigilant, L., M. Stoneking, H. Harpending, K. Hawkes & A.C. Wilson. 1991. African Populations and the Evolution of Human Mitochondrial DNA. Science 253: 1503–7.
Walter, R.C. et al. 2000. Early human occupation of the Red Sea coast of Eritrea during the last interglacial. Nature 405: 65–69.
Weidenreich, F. 1940. Some problems dealing with ancient man. American Anthropologist 42: 380–82.
Wu, X.Z. 1981. A well-preserved cranium of an archaic type of early Homo sapiens from Dali, China. Scientia Sinica 24: 530–41.
Ziętkiewicz, E. et al. 2003. Haplotypes in the Dystrophin DNA Segment Point to a Mosaic Origin of Modern Human Diversity. The American Journal of Human Genetics 73: 994–1015.