{kind=link}
අතීත හෝමෝවරුන් අතරින් වඩාත් ප්රකට සහ වඩාත් මතහේදාත්මක සංවාද සදහා මුල පිරූ මානව විශේෂය වූයේ හෝමෝ නියැන්ඩතාල් ය. නියැන්ඩතාල් මානවයා මුලින් හෝමෝ සේපියන් මානව ගණයට ඇතුළත් වූ නමුත් උප විශේෂ තත්ත්වයෙන් එනම් හෝමෝ සේපියන්ස් නියැන්ඩතාලෙන්සිස් (Homo sapiens neanderthalensis) මගින් වෙන්කර හඳුනාගෙන ඇත. DNA විශ්ලේෂණයෙන් ලැබෙන සාක්ෂි මගින් අදින් වසර ලක්ෂ 3කට පෙර පෙළපත් දෙක බෙදී ගිය බවත්, නව DNA සාක්ෂි මගින් එම දින නිර්ණය කිරීම් තවත් ආපස්සට එනම් අදින් වසර ලක්ෂ 8ක් දක්වා අතීතයකට ඇද දැමිය හැකි බව විද්යාඥයන්ගේ මතයයි. කෙසේ වෙතත්, DNA සාක්ෂි පෙන්නුම් කරන්නේ, නූතන මානවයින් හා නියැන්ඩතාල් මානවයින් අන්තර් අභිජනනය කර ඇති බවට බවත්, සමහරවිට කායච්ඡේද විද්යාත්මකව නූතන මානවයා එක් වරක දී හෝ කිහිප වරකදී අප්රිකාවෙන් පිට වී ගොස්, ඒ වන විටත් මැදපෙරදිග හා යුරෝපයේ ජීවත් වූ නියැන්ඩතාල් මානවයින් සමග සහයෝගයෙන් ජීවත් වූ බවයි. වර්තමානයේ දී වුවද යුරේසියානුවන් සහ ඕස්ට්රේලියානුවන් සාමාන්යයෙන් 2.5% නියැන්ඩතාල් මානව ජාන සංයුතියකින් සමන්විත ය (Noonan 2010). මේ අනුව, කායච්ඡේද විද්යාත්මකව නූතන මානවයාගේ මූලාරම්භය සඳහා අප්රිකානු සම්භවයක් සහිතව ගොඩනගන ලද ආකෘතිය තවමත් වලංගු වන අතර නියැන්ඩතාල් මානවයා අප්රිකාවෙන් පිටත පරිණාමය වූ වෙනම මානව විශේෂයක් ලෙස විද්වතුන් පිළිගනී. නමුත් නියැන්ඩතාල් මානවයා සතු සමහර කායච්ඡේදවිද්යාත්මක ලක්ෂණ හා සංජානන හැකියාවෙන් නූතව මානවයා අභිබවා සිටි බව ද නොරහසකි.
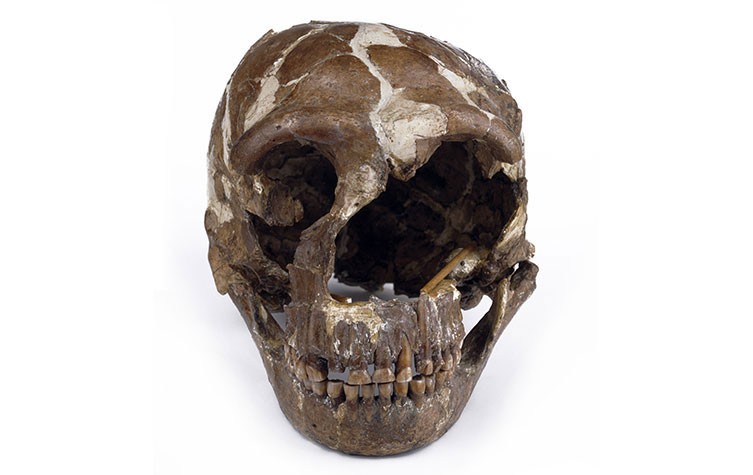
නියැන්ඩතාල් මානවයා සතු සලකුණු අදින් වසර ලක්ෂ 3 සිට 30,000ක් දක්වා පමණ කාලයකට අදාලව බටහිර යුරේසියාවෙන් හමුවේ. නමුත් යුරෝපයේ ප්රථම නූතන මානවයා පෙනීසිටීමෙන් පසු සහස්ර කිහිපයකට පසු ඔවුන් ෆොසිල වාර්තාවෙන් අතුරුදහන් විය (අවම වශයෙන් අදින් වසර 40 000 පෙර). එකී ෆොසිල සුවිශේෂී ව්යුහ විද්යාත්මක ලක්ෂණ වල අද්විතීය සංකලනයකින් සංලක්ෂිත වන අතර මුස්ටේරියන් ශිලා මෙවලම් සමග සන්දර්භ ගත වී දක්නට ලැබේ. වර්තමාන සම්මුතිය ඔවුන්ව පැරණි ලෝකයේ කොටස් වලින් (අප්රිකානු මහද්වීපය) හුදකලා වූ සහ මුල්කාලීන මධ්ය ප්ලයිස්ටොසීන යුගයේ වූ නූතන මිනිසුන් සමඟ පොදු පූර්වජ ලක්ෂණ බෙදාගත් යුරේසියානු මානව පරම්පරාවක් ලෙස සලකයි. යුරෝපයේ වූ අයිස් යුගයේ අධික සීතල, නියැන්ඩතාල් මානවයා සතු වූ කායව්යුහ විද්යාවේ පරිණාමය සඳහා අඩු වැඩි වශයෙන් වගකිව යුතු යැයි සැලකේ.
පළමු නියැන්ඩතාල් ෆොසිල
කායච්ඡේදවිද්යාත්මකව නූතන මානවයා හා නියැන්ඩතාල් මානවයා පිළිබද කතිකාවත සවිගැන්වෙන ප්රධාන පර්යේෂණ ආකාර ද්විත්වයකි. එනම් ෆොසිල සාධක හා ජාන විද්යාත්මක සාධක ලෙස ය. ෆොසිල සාධක කෙරෙහි අවධානය යොමු කළ විට පැහැදිලි වෙන්නේ කායච්ඡේද විද්යාත්මකව නූතන මානවයා හා නියැන්ඩතාල් මානවයා උපත ලබා ඇත්තේ හෝමෝ හයිඩ්ල්බර්ගෙන්සිස් (Homo heidelbergensis) මානවයාගෙන් වන අතර, එම මානවයා අප්රිකානු සම්භවයක් සහිත හෝමෝ අර්ගෙස්ටර් (Homo ergaster) මානවයා ගුරු කොට පරිණාමය වී ඇති බවත් ය. නවතම ජානවිද්යාත්මක සාක්ෂි අනුව කායච්ඡේදවිද්යාත්මකව නූතන මානවයා හා නියැන්ඩතාල් මානවයාගේ පරම්පරා අදින් වසර ලක්ෂ 8කට පමණ පෙර අප්රිකාවේ දී වෙන් වී ඇති බවත්, පසුව එය ඩෙනිසෝවන් මානවයා හා හෝමෝ හයිඩ්ල්බර්ගෙන්සිස් මානවයා දක්වා දිගු වන බවත්, නියැන්ඩතාල් හා ඩෙනිසෝවන් ගහන එකිනෙකින් ජාන විද්යාත්මකව වෙන් වූයේ අදින් වසර ලක්ෂ 6කට පමණ පෙර බවත් තහවුරු වේ. කෙසේ වෙතත්, මෙලෙස දේශීයකරණය වූ කණ්ඩායම් අතර අන්තර් අභිජනනය අඛණ්ඩව සිදු වූ බවට ද ජාන විද්යාත්මක පර්යේෂණ සාක්ෂි දරයි (Rogers et al. 2017).
නියැන්ඩර්තාල් මානව අවශේෂ 19 සියවසේ මුල් භාගයේදී සොයා ගන්නා ලද නමුත් (Engis child 1830, Forbes Quarry adult 1848), වර්ෂ 1856 දී ජර්මනියේ නියැන්ඩර් මිටියාවතෙන් ෆොසිල සැකිල්ල සොයා ගන්නා තෙක් ඒවායේ වැදගත්කම හඳුනා නොගත්තේ ය. එම සොයාගැනීම දළ වශයෙන් 1859 දී ඩාවින්ගේ On The Origin of Species කෘතිය හා සමකාලීන වේ. මෙම විශේෂ මානව ගණය සදහා නියැන්ඩතාල් යන නාමය එහි පළමු ෆොසිල අනාවරණය කරගත් ප්රදේශය ගුරුකොට ගත් අතර, මෙම ෆොසිලවිද්යාත්මත සොයාගැනීම නූතන මිනිසුන් සමඟ නියැන්ඩතාල්වරුන්ගේ සම්බන්ධතාවය වටා අද වන තෙක් සංකේන්ද්රණය වී ඇති අඛණ්ඩ විවාදයේ ආරම්භය සනිටුහන් කරයි.
නමුත්, එලෙස අනාවරණය කරගත් නියැන්ඩතාල් මානව සැකිල්ලේ පෞරාණිකත්වය සහ වඳ වී ගිය මානව විශේෂයක් ලෙස එහි තත්ත්වය වහාම පිළිගැනීමට ලක් නොවිණි. ඒ වෙනුවට, එහි වූ සුවිශේෂී බාහිර රූපවිද්යාත්මක ලක්ෂණල නූතන මිනිස් කණ්ඩායමක් සතුව පැවති විවිධ ව්යාධි (Pathologies) ලෙස ආරෝපණය විය. එහි පෞරාණිකත්වය ස්ථිරව තහවුරු වූයේ, ශිලාමෙවලම් සහ වඳ වී ගිය සතුන්ගේ අස්ථි සමඟ එකට හමු වූ අතිරේක සමාන ෆොසිල සැකිලි සොයා ගැනීමත් සමඟ පමණි. නූතන මිනිසුන්ගේ ෆොසිල පූර්වගාමීන් ලෙස ඔවුන්ගේ පෞරාණිකත්වය පිළිගත් පසු, නූතන මානවයන් සහ විශේෂයෙන්ම නූතන යුරෝපීයයන් සමඟ නියැන්ඩතාල්වරුන්ගේ සබඳතාව දැඩි ලෙස විවාදයට ගැනීමට පටන් ගත්තේය. 1910 – 1920 කාලයේ ප්රමුඛ මතය නියෝජනය කළේ ඔවුන්ගේ කාලයේ සිටි Marcellin Boule සහ Sir Arthur Keith වැනි විද්යාඥයන් විසිනි. ඔවුන් නියැන්ඩතාල් මානවයා වෙනම විශේෂයක් ලෙස හුදකලා කළ අතර, නූතන මිනිසුන්ගේ පරිණාමීය ලක්ෂණ සමග සාපේක්ෂව ගත් කළ නියැන්ඩතාල්වරුන් ප්රක්ෂේපණය කළ ප්රාථමික අස්ථි විද්යාත්මක ලක්ෂණ හේතුවෙන් නූතන මානවයා සමග තිබිය හැකි සියළුම ආකාරයේ පරිණාමීය සබදතා ප්රතික්ෂේප කරන ලදී (Boule 1909).
නියැන්ඩතාල් මානවයා කෙරෙහි වූ මෙම සංජානනය 1930 ගණන්වල සිට වෙනස් විය. ජීව විද්යාවේ නූතන භාවිතය සමග Mayr, Simpson සහ Dobzhansky යන විද්යාඥයින්, නියැන්ඩතාල් මානවයින් සහ අනෙකුත් ප්ලයිස්ටොසීන ෆොසිල මානවයන් නූතන හෝමෝ සේපියන්වරුන්ට නෑකම් දැක්විය හැකි පරිණාමීය කුලකයක් තුළ තැන්පත් කරන ලදී. මෙම මතයට අනුව, නියැන්ඩතාල් මානවයින් මන්දගාමී නමුත්, ක්රමානුකූල පරිණාමය හරහා නූතන මිනිසුන් බවට පරිණාමය වී ඇතැයි සැලකේ (Tattersall 2000; Trinkaus & Shipman 2005). වර්තමානයේ දී පුරාමානවවිද්යාඥයින් බහුතරයක් නියැන්ඩතාල් මානවයා සලකන්නේ වෙනස් බටහිර යුරේසියානු පරිණාමීය පරම්පරාවක් ලෙස වන අතර, එය බොහෝ විට නූතන මිනිසුන්ගේ පරිණාමයට සැලකිය යුතු දායකත්වයක් ලබා නැති බව පිළිගැනේ.
භූගෝලීය පැතිරීම
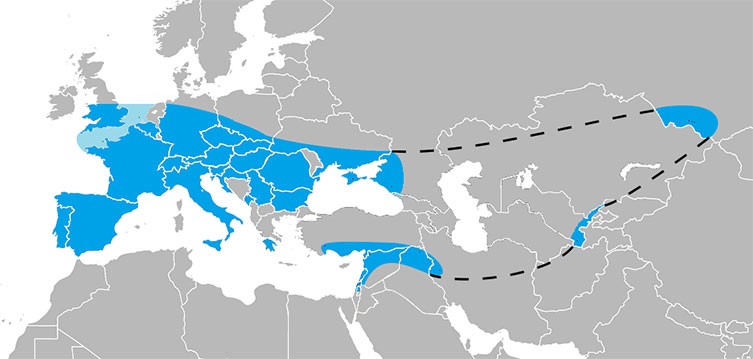
මෙතෙක් යුරෝපයේ හදුනාගෙන ඇති පැරණිතම මානව අවශේෂ ආසන්න වශයෙන් වසර මිලියන 1.2 කට පෙර කාල නිර්ණය කර ඇති අතර ඒවා ස්පාඤ්ඤයේ Sima del Elefante (Carbonell et al. 2008) වෙතින් සොයා ගෙන ඇත. බොහෝවිට හෝමෝ ගණයේ මුල් පුර්වජ ගහන ලෙස හැඳින්වෙන මෙම මුල් යුරෝපීය මානව කොටස, පසුකාලීන, යුරෝපීය, හෝමෝ හයිඩ්ල්බර්ගෙන්සිස් සහ නියැන්ඩතාල් ට උපත ලබා දුන් මූල ලෙස විද්වතුන් සලකයි (Bermúdez de Castro et al. 1997).
ෆොසිල වාර්තාවේ නියැන්ඩතාල්වරුන්ගේ පහළ වීම අතිශයින් ක්රමානුකූල වන අතර, අදින් වසර 600,000 කට පෙර (Bischoff et al. 2003) යුරෝපියානු මානව ෆොසිලවල (හෝමෝ හයිඩ්ල්බර්ගෙන්සිස් ලෙස ද මුල් කාලීනව හැඳින්විණි) නියැන්ඩතාල් ලක්ෂණ ප්රථම වරට දිස්වේ. එකී නියැන්ඩතාල් ලක්ෂණවල තීව්රතාවය කාලයත් සමඟ වැඩි වන අතර, ආසන්න වශයෙන් අදින් වසර 200, 000 සිට 100,000 දක්වා කාලයකට පෙර පටන් නිශ්චිතවම නියැන්ඩතාල් අස්ථිවිද්යාත්මක ලක්ෂණ පෙන්වයි. ආසන්න වශයෙන් අදින් වසර 70,000 සිට 30,000 පමණ පෙරවූ ප්ලයිස්ටොසීන් යුගයේ “සම්භාව්ය”නියැන්ඩර්තාල් ලක්ෂණ නිරීක්ෂණය කළ හැකි ය. නියැන්ඩතාල් වැනි ලක්ෂණ ක්රමක්රමයෙන් තීව්රවෙමින් පෙන්නුම් කරන්නේ කාලයාගේ ඇවෑමෙන් යුරෝපයේ හෝමෝ හයිඩ්ල්බර්ගෙන්සිස් මානවයාගේ සිට නියැන්ඩතාල්වරුන් දක්වා අඛණ්ඩ පරිණාමීය ගමනක් පැමිණි බව ය (Rosas et al. 2006; Tattersall & Schwartz 2006). නියැන්ඩතාල් පරිණාමයේ මෙම ක්රියාවලිය විස්තර කර ඇත්තේ “අධිවර්ධන ආකෘතිය (Accretion Model)” ලෙසිනි (Dean et al. 1998; Hublin 2009).
ආසන්න වශයෙන් අදින් වසර 30,000 කට පමණ පෙර නියැන්ඩතාල්වරු ෆොසිල වාර්තාවෙන් අතුරුදහන් විය, ඒ වසර 40,000 කට පමණ පෙර යුරෝපයේ නූතන මානවයන් පළමු වරට පෙනී සිට සහස්ර කිහිපයකට පසුවය. අයිබීරියාවේ නියැන්ඩර්තාල් මානවයන් අවසන් වරට පෙනී සිටි දිනය ලෙසින් වසර 24,000කට පෙර ඉතා මෑත කාලීන දිනයක් ලබා දී ඇති නමුත් (Finlayson et al. 2006) ස්තරගත පිහිටීම් හා ඇති නොගැලපීම් (Delson & Harvati 2006) හේතුවෙන් එහි වලංගුභාවය ප්රශ්න කිරීමට ලක් වී ඇත.
යුරෝපය තුළ, නියැන්ඩතාල්වරු අයිබීරියාවේ සිට රුසියාව දක්වා සහ මධ්යධරණී මුහුදේ සිට උතුරු යුරෝපය දක්වා තම වාසස්ථාන ලෙස භාවිත කර ඇත. ග්ලැසියර කාලවලදී නියැන්ඩතාල්වරුන් යුරෝපයේ දකුණු අර්ධද්වීපයට සංක්රමණය වී ඇති අතර (Harvati et al. 2003, 2009; Finlayson et al. 2006), යුරෝපයේ දැඩි දේශගුණික සීමා මායිම්වලින් පිටත, නියැන්ඩතාල් මානව වාසස්ථාන පෙරදිග සහ බටහිර ආසියාවේ උස්බෙකිස්තානය සහ සයිබීරියාවේ පවා අනාවරණය වී ඇත (Krause et al. 2007).
කායික විද්යාත්මක ලක්ෂණ
නියැන්ඩතාල් මානවයින් බොහෝ විට ඔවුන්ටම ආවේණික කායික විද්යාත්මක ලක්ෂණවලින් සමන්විත ය. එනම්, හිස්කබල, දන්ත වින්යාසය හා පශ්චාත් හිස්කබල ආශ්රිත ලක්ෂණ රාශියකින් සංලක්ෂිත ය. ඊට අමතරව ප්රාථමික යැයි හැදින්විය හැකි ලක්ෂණ කීපයක් ද ඔවුට පෙර සිටි පූර්වජ ගහන සමග බෙදාහදා ගත් බවක් ද දක්නට ලැබේ (Harvati 2015). තවදුරටත් නියැන්ඩතාල්වරුන්ගේ මුහුණ බර, අඛණ්ඩ සහ ද්විත්ව ආරුක්කු සහිත අස්ථි හා ඇහි බැම මගින් අනෙකුත් මානවයින්ගෙන් වෙන්කර හදුනාගත හැකි වේ. අක්ෂි කුහර විශාල හා වටකුරු ස්වභාවයක් ගනී. නාසික විවරය ඉතා විශාල හා පුළුල් වන අතර, මධ්යම මුහුණේ මදක් වැඩුණු අස්ථිමය ප්රක්ෂේපණයක් සහිත ය, කම්මුල් අස්ථි වක්ර ලෙස දිශානත වී ඇත. දන්ත වින්යාසය සැලකූ විට යටි හකු පසු බසිමින් පවතින බවක් නිරීක්ෂණය කෙරෙන අතර, නෙරා ඇති නිකටක් නොමැත; තවද, පහළ චාර්වක දත් සහ යටි හකු අතර පැහැදිලි ඉඩක් ඇත (Retromolar space) (Trinkaus 1987; Franciscus 2003; Nicholson & Harvati 2006). නියැන්ඩතාල් මානව පසුපස දත්වල විහිදීම, සම්පුර්ණයෙන්ම නූතන මිනිසුන්ගේ දත් සමඟ අතිච්ඡාදනය වේ. කෙසේ වෙතත්, නියැන්ඩතාල් මානව ඉදිරිපස දත් සහ විශේෂයෙන් කෘන්තක විශාල වන අතර නූතන මිනිසුන්ට සාපේක්ෂව දන්ත ව්යුහ විද්යාත්මක ලක්ෂණ කිහිපයක් නියැන්ඩර්තාල් මානවයන් තුළ බහුලව දක්නට ලැබේ (Bailey 2002, 2004). විශේෂ කරුණක් ලෙස නියැන්ඩතාල් මානව දත් ගෙවීයාමේ විශේෂ රටාවකින් නිරීක්ෂණය වේ, එනම්, ඉදිරිපස දත් සාමාන්යයෙන් එකම පුද්ගලයාගේ පිටුපස දත් වලට වඩා බොහෝ සෙයින් ගෙවී ගොස් ඇත (Keith 1913; Trinkaus 1983).

නියැන්ඩතාල් මානවයා සතු විශාල නාස් කුහරය සහ ආශ්රිත ව්යුහ ශීත දේශගුණයට අනුවර්තනය වීමට බොහෝ කලක සිට අනුවර්තනය වී අතර විද්වත් මතය අනුව, සීත වාතය උණුසුම් කිරීම උෂ්ණත්වය යාමනය කිරීම සඳහා හැඩගැසී ඇති බවයි (Dean 1988; Hubbe et al. 2009). එමෙන්ම ඔවුන්ගේ අසමමිතික දන්ත රටාව ඔවුන් දත් කාර්යකරන මෙවලම් ලෙස භාවිත කිරීම තුළින් සිදුවූ ජෛව යාන්ත්රික ප්රතිවිපාක බව විද්වත් මතයයි (Trinkaus 1983; Rak 1986). තුන්වන අර්ථකථනය නම්, නියැන්ඩතාල් මානවයාගේ රූප විද්යාව මූලික වශයෙන් අහඹු ලෙස මෙහෙයවන ක්රියාවලීන්ගේ (එනම් ජාන ප්ලාවිතය වැනි) ප්රතිඵලයක් ලෙස සැලකීමයි (Dean et al. 1998; Weaver et al. 2007).
නියැන්ඩතාල් මානවයාගේ සාමාන්ය කපාල ධාරිතාවය නූතන මිනිසුන්ට වඩා විශාල වන අතර එය ආසන්න වශයෙන් ඝන සෙන්ටිමීටර 1520 ක (ඝන සෙන්ටිමීටර 1,200 සිට 1,700 දක්වා) පරාසක් ගනී (Pearce et al. 2013). නියැන්ඩතාල් මානවයාගේ කපාල ධාරිතාවය සීතල දේශගුණයට අනුවර්තනය වීමට පිටිවහලක් වූ බවට ද අදහස් පල වී ඇත (Churchill 1998). නූතන මිනිසුන්ට සාපේක්ෂව නියැන්ඩතාල් මානවයාගේ කපාල ධාරිතාවය විශාල වුවද, එය ඔවුන්ගේ විශාල ශරීර ස්කන්ධයට සාපේක්ෂව කුඩා විය හැකිය (Ruff et al. 1997). නියැන්ඩතාල් සහ නූතන මානව යන දෙපාර්ශවයේ ම කපාල ධාරිතාවය විශාල වීමේ ලක්ෂණය පරම්පරා දෙකෙහි දක්නට ලැබෙන වෙනස් පරිණාමීය ගමන් පථ අනුගමනය කිරීමේ ප්රතිඵලයක් බව නිසැක ය (Bruner et al. 2003).
නියැන්ඩතාල් සැකිල්ල සමස්තයක් ලෙස ශක්තිමත් වන අතර, කලවා සහ අත්වල, ඝන අස්ථි, සහ ශක්තිමත් මාංශ පේශී සහ අස්ථිබන්ධන (ligament) සලකුණු සහිත ව කැපී පෙනේ. ඇස්තමේන්තුගත උස සාමාන්යයෙන් පිරිමින් සඳහා සෙන්ටිමීටර 169 ක් සහ කාන්තාවන් සඳහා සෙන්ටිමීටර 160 ක් පමණ වන අතර එය මුල් මානව පූර වජයන්ට වඩා අඩු අගයකි. තවදුරටත් ඔවුන්ගේ ඉළ ඇට (rib cage) පුළුල් හා ගැඹුරු වන අතර ඉහළ ප්රදේශයෙන් පටු ආකාරයක් ගන්නා අතර, (Franciscus & Churchill 2002; Sawyer & Maley 2005) සාපේක්ෂව කෙටි දුරස්ථ අත් පා සහ ජංඝක අස්ථිය (Tibia) සහ ඌර්වස්ථීය (Femur) කෙළවර ඝන ලෙස නිමවී තිබිණි (බොහෝ විට සීත දේශගුණයට දක්වන ලද කායච්ඡේදවිද්යාත්මක අනුවර්තනයක් ලෙස) (Trinkaus 1983; Ruff 1991; Ruff et al. 1997; Weaver 2003). තවදුරටත්, නියැන්ඩතාල් මානව කලවයේ සහ ශ්රෝණියෙහි ඇති ශක්තිමත් බව හා පුළුල් බව වැනි ලක්ෂණ ද ශීත දේශගුණයට සම්බන්ධ ලෙස විද්වතුන් විසින් අර්ථකථනය කර ඇත (Pearson 2000; Weaver 2003). නමුත් මෙම අනුවර්තන නියැන්ඩතාල් මානවයා අදාල පරිසර තත්ත්ව තුළ නොනැසී රැකීමේ ක්රියාන්විතයේ දී අවාසිදායක ලෙස සටහන් වී ඇත (Aiello & Wheeler 2003).
මෙතෙක් සිදු කර ඇති අධ්යයන අනුව නියැන්ඩතාල් දරුවන් නූතන සේපියන් දරුවන්ට වඩා වේගයෙන් වර්ධනය වූ බව අනාවරණය වී ඇත (Dean et al. 2001; Ramirez Rozzi & Bermudez De Castro 2004; Smith et al. 2007). නියැන්ඩතාල්වරුන්ට ද වෙනස් ජාන විකාශනයන් (demography ) ඇති බව පෙනේ: නියැන්ඩතාල් ක්ෂේත්ර තුලින් නව යොවුන් වියේ සහ ප්රධාන වයස්වල (prime age adults) වැඩිහිටියන්ගේ ප්රතිශතය වයොවෘධ වැඩිහිටියන් හා ළදරුවන්ට සාපේක්ෂව අසාමාන්ය ලෙස ඉහළ ය. මෙම රටාව අනුරූපීයව අඩු වැඩිහිටි ආයු අපේක්ෂාවක් (low adult life expectancy) හා තරුණ හා ප්රමුඛ වයස්වල නියැන්ඩතාල්වරුන් අතර ඉහළ මරණ අනුපාතිකයක් (high mortality rate) යෝජනා කරයි (Berger & Trinkaus 1995; Trinkaus 2011).
තාක්ෂණය, ආහාර සහ හැසිරීම් රටාව
ප්රංශයේ Dordogne හි Le Moustier නම් ස්ථානයෙන් හමු වී ඇති අවශේෂ මත නම් කර ඇති මුස්ටේරියන් ශිලාමෙවලම් තාක්ෂණය සමඟ නියැන්ඩතාල්වරු ඍජුවම සම්බන්ධ වේ. මුස්ටේරියන් (Mousterian) ශිලාමෙවලම් සාමාන්යයෙන් පුළුල් පරාසයක හැඩතල සහ කාර් යය මෙවලම් බවට පරිවර්තනය කළ හැකි ශිලා පතුරු නිෂ්පාදනය සඳහා ලෙවාලුවා (Levallois) සහ මණ්ඩලාකාර පතුරු ගැසීමේ (discoidal flaking) ශිල්පීය ක්රම උපයෝගී කරගෙන ඇත. මුස්ටේරියන් මෙවලම් යුරෝපයේ වසර 200,000 සිට 150,000 දක්වා කාලයකට පෙර අනාවරණය වූ නමුත්, බොහෝ ක්ෂේත්ර වසර 130,000 සිට 30,000 දක්වා කාල පරාසයක් දක්වා පැරණි ය. බොහෝ මුස්ටේරියන් ක්ෂේත්රවල ශිලාමෙවලම් නිෂ්පාදනය සඳහා භාවිතා කරන අමුද්රව්ය එම ස්ථානයේ සිට කිලෝමීටර් පහක සිට හය දක්වා පරාසයකින් ව්යුත්පන්න කරගැනීමට නැඹුරු වී ඇති අතර කුඩා සංරචක ප්රමාණයක් පමණක් දුරස්ථ මූලාශ්රවලින් සම්පාදනය කරගනෙ ඇත. ජර්මනියේ ෂනිගර් (Schöningen) හි සොයා ගන්නා ලද සහ ආසන්න වශයෙන් වසර 400,000කට පෙර දින නියම කරන ලද හොඳින් සංරක්ෂණය වී ඇති ලී හෙලි කිහිපයකින් සනාථ වන පරිදි, ලී මෙවලම් ද නියැන්ඩතාල්වරුන් භාවිත කර ඇත (Thieme 1997; Shea & Brooks 2000).

මෑතක් වනතුරු, මුස්ටේරියන් ශිලාමෙවලම් ස්ථිතික මානව සංස්කෘතියක් නියෝජනය කරන බව සලකනු ලැබීය. කෙසේ වෙතත්, මුස්ටේරියන් ක්ෂේත්ර නැවත කාලනිර්ණය කිරීම තුළින් කලාපීය මෙලවම් භාවිතය කාලයත් සමඟ මෙන්ම, දේශගුණික විපර්යාස සඳහා ද අනුවර්තිත තාක්ෂණික ප්රතිචාර පෙන්නුම් කරන බව විද්වත් මතයයි (Kuhn 1995).
නියැන්ඩතාල් ක්ෂේත්ර ආශ්රිතව විවිධ සතුන්ගේ සත්ත්ව අවශේෂ බහුලව දක්නට ලැබෙන අතර, ඔවුන්ගේ ආහාර වේලෙහි අධික ලෙස සත්ත්ව මාංශමත රඳා පැවති බව පෙන්නුම් කරයි. බයිසන් ගවයින්, ගවයන්, අශ්වයන්, මුවන්, රතු මුවන්, අයිබෙක්ස්, වල් ඌරා සහ ගැසල් වැනි සතුන් ඒ අතර ප්රධාන වේ (Shea & Brooks 2000). ගොඩබිම් වෙසෙන සතුන් පමණක් නොව බෙල්ලන් හා සාගර ක්ෂීරපායීන් වැනි සතුන් ද, ළගා විය නොහැකි සතුන් ලෙස ගැනෙන කුරුල්ලන් ද නියැන්ඩතාල් මිනිසුන් පරිභෝජනය කර ඇති බවට මධ්යධරණි ප්රදේශයෙන් සාක්ෂි හමුවේ (Hockett & Haws 2005; Stringer et al. 2008a; b; Muñiz et al. 2019).
නියැන්ඩතාල් ෆොසිලවල රසායනික සංයුතිය අධ්යයනය කිරීමෙන් ඔවුන්ගේ ආහාර රටාව හදුනාගැනීමට හැකි ය. නියැන්ඩර්තාල් නිදර්ශක ගණනාවක් ආශ්රයෙන් අස්ථිවල කාබන් සහ නයිට්රජන් වල ස්ථායී සමස්ථානිකවල අනුපාත විශ්ලේෂණය පුළුල් කාල පරාසයක් (වසර 130,000 සිට 30,000 පමණ පෙර) දක්වා සිදු කර ඇත. එවැනි විශ්ලේෂණ ශාකභක්ෂක සත්ත්ව මාංශ මත දැඩි ලෙස රඳා පවතින බව නිරන්තරයෙන් පෙන්වා දී ඇත, තවදුරටත් නියැන්ඩතාල් අස්ථිවල සමස්ථානික සංයුතිය අනුව ඔවුන් ආහාර දාමයේ ඉහළම විලෝපිකයන්ට සමාන වන බව විද්වත් පිළිගැනීමයි (Fizet et al. 1995; Bocherens et al. 1999, 2005; Richards et al. 2000). තවත් අධ්යයනවලින් රයිනෝසිරස් හෝ මැමත් වැනි විශාල සතුන් පරිභෝජනය කිරීම ද යෝජනා කර ඇත (Bocherens et al. 2005). සමස්ථානික විශ්ලේෂණ පර්යේෂණ තුළින් ඉහළ පැලියොලිතික් නූතන මානවයන්ට වඩා තියුනු ලෙස හා වෙනස්ව, නියැන්ඩතාල් මානවයින් තම ආහාර වේල සදහා සමුද්රාශිත ආහාර එක් කරගත් බව සනාථ වේ (Richards et al. 2001, 2005).
නියැන්ඩතාල්වරුන්ගේ සංකේතාත්මක චින්තනය සහ භාෂාව සඳහා වූ හැකියාව තවමත් පර්යේෂණ මට්ටමේ පවතී. කලාත්මක හෝ පුද්ගලික ආභරණ වැනි හැකියාවන් යැයි සැකකළ හැකි “සංකේතාත්මක” පුරාකෘති, පශ්චාත් ඉහළ පැලියොලිතික් ස්ථාන සමඟ පමණක් නොව, නියැන්ඩතාල් ක්ෂේත්ර ආශ්රිතව ද අතිශයින් දුර්ලභ වේ. එවැනි සාක්ෂි නොමැතිකම මානව සංජානන හැකියා සහ භාෂාවේ ඌනතාවයක් පෙන්නුම් කිරීම උදෙසා භාවිතා වී ඇත. නමුත් නියැන්ඩතාල්වරුන් මියගිය පසු භුමදානය සිදු කිරීම හා මියගියඅය සදහා මල් පිදීම ඔවුන් ලබා සිටි ඉහළ සංස්කෘතික චාරවත් බව ප්රකට කරයි. ඒ අනුව නියැන්ඩතාල්වරු යනු ඔවුන්ගේ මළවුන් තනිව සොහොන් වල මිහිදන් කළ (individual graves) පළමු විශේෂයයි. සිරුරු බොහෝ විට නවා ඇත (flexed position). ඔවුන් පොළොවේ වළක් හාරා, ශරීරය ඒ තුළට නමා, සමහර විට ඒ සමඟ තවත් අයිතම නිදන් කළ බවට සාක්ෂි හමුවේ. එම ද්රව්ය බොහෝ විට යම් පුද්ගලික වැදගත්කමක් ඇති ද්රව්ය ලෙස අර්ථකථනය කරනු ලැබේ, ඒ අතර සත්ව ඇටකටු සහ කැඩුණු මෙවලම් බහුල ව දක්නට ලැබේ.

නියැන්ඩතාල් මානවයා ලෝකයෙන් වද වී යාම
බොහෝ විද්වතුන්ගේ මතය වන්නේ, නියැන්ඩතාල්වරුන් අවසානයේ, ඔවුන් නූතන යුරෝපීයයන් බවට පරිණාමය වීම හෝ නූතන මානව ජාන සංචිතය විසින් ඔවුන්ව අවශෝෂණය කරගත් බවයි. කෙසේවෙතත්, නූතන මිනිසුන් සමඟ සීමිත ජාන හුවමාරුවක් පමණක් සිදු වුවද, බොහෝ දෙනෙක් එය සැබෑ වඳවීමක් ලෙස සලකති. නියැන්ඩතාල්වරුන්ගේ වඳවීම සඳහා උපන්යාස කිහිපයක් යෝජනා වී ඇති අතර, බොහෝ විට ඒවා මුල් නූතන මානවයන් සමඟ ඇති වූ සෘජු හෝ වක්ර තරඟකාරීත්වයකින් පසුව සිදු වී බව කියයි. සමහර විද්යාඥයින් යෝජනා කරන්නේ ඔවුන්ගේ වඳවීම පසුකාලීන ප්ලයිස්ටොසීන් දේශගුණ විපර්යාසයත් සමග සමකාලීන වන බවයි (Stewart 2005). පුරාමානවවිද්යාඥයන් වෙනස් වූ දේශගුණික සහ පාරිසරික තත්ත්වයන් නියැන්ඩතාල්වරුන් වඳවීමේ ප්රධාන ගාමක බලවේගයන් ලෙස සලකයි. නියැන්ඩතාල්වරුන් ෆොසිල වාර්තාවෙන් අතුරුදහන් වූ කාල සීමාව ඉතා අස්ථායී දේශගුණික තත්ත්වයන් විසින් ආධිපත්යය දැරූ බව මෑතකදී ලබා ගත හැකි සවිස්තරාත්මක පුරාකාලගුණවිද්යාත්මක වාර්තා මගින් පෙන්වා දී ඇත (Straus 2005). නියැන්ඩතාල් මානවයන් පෙර පැවති සීතල දේශගුණය සදහා මූලිකව අනුවර්තනය වුවද, ඔවුන්ගේ වද වීමට එකම හේතුව ලෙස දේශගුණික විපර්යාස පිළිගැනීම දුෂ්කර වන්නේ නූතන මානවයාට වඩා වැඩි කපාල ධාරිතාවකින් හෙබි, පරිසරය පිළිබද අනල්ප වූ දැනුමක් තිබූ මානව විශේෂයක් සදහා පවත්නා පරිසරයට උචිතව හැඩගැසීමට නොහැකි වීම සාධාරණ හේතුවක් නොවන නිසාවෙනි. විකල්පයක් ලෙස දකින්නේ නූතන මිනිසුන්ගේ ආගමනය සමග වේගවත් වූ දැඩි ලෙස සීමිත සම්පත් සඳහා තරඟයක් පැවතීම තුළින් දැඩි ආතතියට පත් නියැන්ඩතාල් ගහනය ක්රමයෙන් යුරෝපයෙන් මැකී ගිය බව ය. ඒ බව ඔවුන්ගේ ශිලා මෙවලම් ආශ්රයෙන්, එනම් දියුණු තාක්ෂණික අභිවර්ධනයක් පෙන්නුම් නොකරන ආයුධ කට්ටල මගින්, තාක්ෂණික වශයෙන් දියුණුවේ හිනිපෙත්තටම ගොස් සිටි හෝමෝ සේපියන් මානවයා සමග තරග කළ නොහැකි විය (Bowler et al. 2003; Harvati 2015; Wren & Burke 2019). ඒ අනුව, උච්චාවචනය වූ දේශගුණය සහ පරිසරයේ බලපෑම් සහ නූතන මිනිසුන් සමඟ තරඟකාරිත්වය අතර අන්තර් ක්රියාකාරිත්වය අවසානයේ නියැන්ඩතාල්වරුන්ගේ අභාවයට හේතු වූ බව වර්තමානයේ පිළිගන්නා මතය වී තිබේ.
කෙටි වාර්තා චිත්රපට
- Who were the Neanderthals? | DW Documentary
- Neanderthal – Ancient Human
- Neanderthals Human Extinction BBC Documentary
- Neanderthals 101 | National Geographic
- A Day In The Life Of A Neanderthal
මනුෂ්යත්වයේ කතාන්දරය: මානව පරිණාමය
- මානව පරිණාමයේ ගමන 1: මනුෂ්යත්වයේ ආරම්භය
- මානව පරිණාමයේ ගමන 2- මනුෂ්යත්වයේ මූල බීජ: මුල් වානර මානවයින් හා ඔස්ට්රලෝපිතිකස් ගණය
- මානව පරිණාමයේ ගමන 3- මුල් හෝමෝ සාමාජිකයින්: හෝමෝ හැබිලිස් හා හෝමෝ ඉරෙක්ටස්
- මානව පරිණාමයේ ගමන 4 – හෝමෝ නියැන්ඩතාල් මානවයා
- හෝමෝ සේපියන්ස් මානවයා
- ඩෙනිසෝවන් මානවයා හා මානව පරිණාමයේ නව මානව විශේෂ
- මානවයා අප්රිකාවෙන් පිටවී යාම
- මානවයා නවලොව ජනාවාසකරණය කිරීම (අමරිකාව හා ඕස්ට්රේලියාව)
- නියෝලිතික විප්ලවය හා මුල් කෘෂිකර්මයේ නැගීම
References
Aiello, L. & P. Wheeler. 2003. Neanderthal thermoregulation and the glacial climate, in T. van Andel & W. Davies (ed.) Neanderthals and modern humans in the European landscape during the last glaciation: 147–66. Cambridge: McDonald Institute for Archaeological Research.
Bailey, S.E. 2002. A closer look at Neanderthal postcanine dental morphology: The mandibular dentition. The Anatomical Record 269: 148–56.
Bailey, S.E. 2004. A morphometric analysis of maxillary molar crowns of Middle-Late Pleistocene hominins. Journal of human evolution 47: 183–98.
Berger, T.D. & E. Trinkaus. 1995. Patterns of Trauma among the Neandertals. Journal of Archaeological Science 22: 841–52.
Bermúdez de Castro, J.M., J.L. Arsuaga, E. Carbonell, A. Rosas, I. Martı́nez & M. Mosquera. 1997. A Hominid from the Lower Pleistocene of Atapuerca, Spain: Possible Ancestor to Neandertals and Modern Humans. Science 276: 1392–95.
Bischoff, J.L., D.D. Shamp, A. Aramburu, J.L. Arsuaga, E. Carbonell & J.M. Bermudez de Castro. 2003. The Sima de los Huesos Hominids Date to Beyond U/Th Equilibrium (>350kyr) and Perhaps to 400–500kyr: New Radiometric Dates. Journal of Archaeological Science 30: 275–80.
Bocherens, H., D. Billiou, A. Mariotti, M. Patou-Mathis, M. Otte, D. Bonjean & M. Toussaint. 1999. Palaeoenvironmental and Palaeodietary Implications of Isotopic Biogeochemistry of Last Interglacial Neanderthal and Mammal Bones in Scladina Cave (Belgium). Journal of Archaeological Science 26: 599–607.
Bocherens, H., D.G. Drucker, D. Billiou, M. Patou-Mathis & B. Vandermeersch. 2005. Isotopic evidence for diet and subsistence pattern of the Saint-Césaire I Neanderthal: review and use of a multi-source mixing model. Journal of human evolution 49: 71–87.
Boule, M. 1909. L’homme fossile de la Chapelleaux-Saints. Paris.
Bowler, J.M., H. Johnston, J.M. Olley, J.R. Prescott, R.G. Roberts, W. Shawcross & N.A. Spooner. 2003. New ages for human occupation and climatic change at Lake Mungo, Australia. Nature 421: 837–40.
Bruner, E., G. Manzi & J.L. Arsuaga. 2003. Encephalization and allometric trajectories in the genus Homo : Evidence from the Neandertal and modern lineages. Proceedings of the National Academy of Sciences 100: 15335–40.
Carbonell, E. et al. 2008. The first hominin of Europe. Nature 452: 465–69.
Churchill, S.E. 1998. Cold adaptation, heterochrony, and neandertals. Evolutionary Anthropology: Issues, News, and Reviews 7: 46–60.
Dean, C., M.G. Leakey, D. Reid, F. Schrenk, G.T. Schwartz, C. Stringer & A. Walker. 2001. Growth processes in teeth distinguish modern humans from Homo erectus and earlier hominins. Nature 414: 628–31.
Dean, D., J.-J. Hublin, R. Holloway & R. Ziegler. 1998. On the phylogenetic position of the pre-Neandertal specimen from Reilingen, Germany. Journal of Human Evolution 34: 485–508.
Dean, M.C. 1988. Another look at the nose and the functional significance of the face and nasal mucous membrane for cooling the brain in fossil hominids. Journal of Human Evolution 17: 715–18.
Delson, E. & K. Harvati. 2006. Return of the last Neanderthal. Nature 443: 762–63. .
Finlayson, C. et al. 2006. Late survival of Neanderthals at the southernmost extreme of Europe. Nature 443: 850–53.
Fizet, M., A. Mariotti, H. Bocherens, B. Lange-Badré, B. Vandermeersch, J.P. Borel & G. Bellon. 1995. Effect of diet, physiology and climate on carbon and nitrogen stable isotopes of collagen in a late pleistocene anthropic palaeoecosystem: Marillac, Charente, France. Journal of Archaeological Science 22: 67–79.
Franciscus, R.G. 2003. Internal nasal floor configuration in Homo with special reference to the evolution of Neandertal facial form. Journal of human evolution 44: 701–29.
Franciscus, R.G. & S.E. Churchill. 2002. The costal skeleton of Shanidar 3 and a reappraisal of Neandertal thoracic morphology. Journal of human evolution 42: 303–56.
Harvati, K. 2015. Neanderthals and Their Contemporaries, in Handbook of Paleoanthropology: 2243–79. Berlin, Heidelberg: Springer Berlin Heidelberg.
Harvati, K., E. Panagopoulou & P. Karkanas. 2003. First Neanderthal remains from Greece: the evidence from Lakonis. Journal of human evolution 45: 465–73.
Harvati, K., E. Panagopoulou & C. Runnels. 2009. The paleoanthropology of Greece. Evolutionary Anthropology: Issues, News, and Reviews 18: 131–43.
Hockett, B. & J.A. Haws. 2005. Nutritional ecology and the human demography of Neandertal extinction. Quaternary International 137: 21–34.
Hubbe, M., T. Hanihara & K. Harvati. 2009. Climate signatures in the morphological differentiation of worldwide modern human populations. Anatomical record (Hoboken, N.J. : 2007) 292: 1720–33.
Hublin, J.J. 2009. The origin of Neandertals. Proceedings of the National Academy of Sciences 106: 16022–27.
Keith, A. 1913. Problems relating to the Teeth of the Earlier Forms of Prehistoric Man. Proceedings of the Royal Society of Medicine 6: 103–24.
Krause, J. et al. 2007. The derived FOXP2 variant of modern humans was shared with Neandertals. Current biology : CB 17: 1908–12.
Kuhn, S.L. 1995. Mousterian lithic technology: an ecological perspective. Princeton: Princeton University Press.
Muñiz, F. et al. 2019. Following the last Neanderthals: Mammal tracks in Late Pleistocene coastal dunes of Gibraltar (S Iberian Peninsula). Quaternary Science Reviews 217: 297–309.
Nicholson, E. & K. Harvati. 2006. Quantitative analysis of human mandibular shape using three-dimensional geometric morphometrics. American journal of physical anthropology 131: 368–83.
Noonan, J.P. 2010. Neanderthal genomics and the evolution of modern humans: Figure 1. Genome Research 20: 547–53.
Pearce, E., C. Stringer & R.I.M. Dunbar. 2013. New insights into differences in brain organization between Neanderthals and anatomically modern humans. Proceedings of the Royal Society B: Biological Sciences 280: 20130168.
Pearson, O.M. 2000. Postcranial remains and the origin of modern humans. Evolutionary Anthropology: Issues, News, and Reviews 9: 229–47.
Rak, Y. 1986. The Neanderthal: A new look at an old face. Journal of Human Evolution 15: 151–64.
Ramirez Rozzi, F. v & J.M. Bermudez De Castro. 2004. Surprisingly rapid growth in Neanderthals. Nature 428: 936–39.
Richards, M.P., P.B. Pettitt, E. Trinkaus, F.H. Smith, M. Paunović & I. Karavanić. 2000. Neanderthal diet at Vindija and Neanderthal predation: The evidence from stable isotopes. Proceedings of the National Academy of Sciences 97: 7663–66.
Richards, M.P., P.B. Pettitt, M.C. Stiner & E. Trinkaus. 2001. Stable isotope evidence for increasing dietary breadth in the European mid-Upper Paleolithic. Proceedings of the National Academy of Sciences 98: 6528–32.
Richards, M.P., R. Jacobi, J. Cook, P.B. Pettitt & C.B. Stringer. 2005. Isotope evidence for the intensive use of marine foods by Late Upper Palaeolithic humans. Journal of Human Evolution 49: 390–94.
Rogers, A.R., R.J. Bohlender & C.D. Huff. 2017. Early history of Neanderthals and Denisovans. Proceedings of the National Academy of Sciences 114: 9859–63.
Rosas, A., M. Bastir, C. Martínez-maza, A. García-Tabernero & C. Lalueza-Fox. 2006. Inquiries into Neanderthal craniofacial development and evolution: “accretion” versus “organismic” models, in : 37–69.
Ruff, C.B. 1991. Climate and body shape in hominid evolution. Journal of Human Evolution 21: 81–105.
Ruff, C.B., E. Trinkaus & T.W. Holliday. 1997. Body mass and encephalization in Pleistocene Homo. Nature 387: 173–76.
Sawyer, G.J. & B. Maley. 2005. Neanderthal reconstructed. The Anatomical Record Part B: The New Anatomist 283B: 23–31.
Shea, J.S. & A.S. Brooks. 2000. Mousterian, in E. Delson, I. Tattersall, J.A. van Couvering & A.S. Brooks (ed.) Encyclopedia of human evolution and prehistory, 2nd ed.: 453–60. New York: Garland.
Smith, T.M., M. Toussaint, D.J. Reid, A.J. Olejniczak & J.-J. Hublin. 2007. Rapid dental development in a Middle Paleolithic Belgian Neanderthal. Proceedings of the National Academy of Sciences 104: 20220–25.
Stewart, J.R. 2005. The ecology and adaptation of Neanderthals during the non-analogue environment of Oxygen Isotope Stage 3. Quaternary International 137: 35–46.
Straus, L.G. 2005. Neanderthals and Modern Humans in the European Landscape during the Last Glaciation: Archaeological Results of the Stage 3 Project . Tjeerd H. van Andel , William Davies. Journal of Anthropological Research 61: 278–81.
Stringer, C.B. et al. 2008a. Neanderthal exploitation of marine mammals in Gibraltar. Proceedings of the National Academy of Sciences 105: 14319–24.
—. 2008b. Neanderthal exploitation of marine mammals in Gibraltar. Proceedings of the National Academy of Sciences 105: 14319–24.
Tattersall, I. 2000. Paleoanthropology: The last half-century. Evolutionary Anthropology: Issues, News, and Reviews 9: 2–16.
Tattersall, I. & J.H. Schwartz. 2006. The distinctiveness and systematic context of Homo neanderthalensis, in : 9–22.
Trinkaus, E. 1983. The Shanidar Neandertals. Elsevier.
Thieme, H. 1997. Lower Palaeolithic hunting spears from Germany. Nature 385: 807–10.
Trinkaus, E. 1987. The Neandertal face: evolutionary and functional perspectives on a recent hominid face. Journal of Human Evolution 16: 429–43.
—. 2011. Late Pleistocene adult mortality patterns and modern human establishment. Proceedings of the National Academy of Sciences 108: 1267–71.
Trinkaus, E. & P. Shipman. 2005. Neandertals: Images of ourselves. Evolutionary Anthropology: Issues, News, and Reviews 1: 194–201.
Weaver, T.D. 2003. The shape of the Neandertal femur is primarily the consequence of a hyperpolar body form. Proceedings of the National Academy of Sciences 100: 6926–29.
Weaver, T.D., C.C. Roseman & C.B. Stringer. 2007. Were neandertal and modern human cranial differences produced by natural selection or genetic drift? Journal of human evolution 53: 135–45.
Wren, C.D. & A. Burke. 2019. Habitat suitability and the genetic structure of human populations during the Last Glacial Maximum (LGM) in Western Europe. PLOS ONE 14: 12-26.
ela
blog kiyawanna puluvn syndi ekk https://sithuvili.net/