{kind=link}
හෝමෝ හැබිලිස් මානවයා පරිණාමයට පෙර, පළමු වසර මිලියන හතරක් හෝ ඊට වැඩි කාලයක් සඳහා, ෆොසිල වාර්තාව කායච්ඡේදවිද්යාත්මක ලක්ෂණ අතින් ප්රාථමික බව, එනම්, රදනක දතේ ප්රමාණය අඩුවීම, පශ්චාත් හිස්කබලේ (postcranial) වූ දිගටි හැඩය අඩු වීම වැනි කාරණා මගින් සංලක්ෂිත වේ: සහෙලන්ත්රොපස් (Sahelanthropus), ඔරියොන් (Orrorin), අරිඩිපිතෙකස් (Ardipithecus), ඔස්ට්රලෝපිතෙකස් (Australopithecus) සහ පරන්ත්රොපස් (Paranthropus) යන වානර මානව විශේෂ ඒ සදහා උදාහරණ ලෙස ගිණිය හැකි ය. ප්ලයෝසීන යුගය අවසන් වී වසර මිලියන 2.5 කට පමණ පෙර ගෝලීය දේශගුණය වෙනස් වෙමින් පැවතීම නිසා (deMenocal 2004), මානව ෆොසිල වාර්තාවේ සමගාමී වෙනසක් නිරීක්ෂණය කළ හැකිය. පරිසරය වැඩිවශයෙන් සිසිල් වීමක් සමග ෆොසිල වාර්තාවේ කායව්යුහවිද්යාත්මක හා චර්යාත්මකව වෙනස් ලක්ෂණ නිරීක්ෂණය කළ හැකි විය. එය වර්තමාන මානවයා අයත්වන හෝමෝ ගණයේ මූලාරම්භයයි.
මානව ෆොසිල සොයායාම…
1964 දී, ලුවිස් ලීකී (Louis Leakey), පිලිප් ටොබායස් (Philip Tobias), සහ ජෝන් නේපියර් (John Napier) ඔස්ට්රලෝපිතෙකස් දිගුව අවසන් වන්නේ කොතැනින්ද සහ හෝමෝ ආරම්භ වන්නේද යන ගැටළුව වෙත මුල්ම ප්රවිශ්ඨ වීම සනිටුහන් කළේ ය (Leakey et al. 1964). මීට වසර මිලියන 1.75 කට පමණ කාලනිර්ණ කර ඇති, Olduvai Gorge ආශ්රිත හිස් කබලේ කොටස්, හකු කොටස් සහ මැණික් කටුව සහ ගාත්රා අස්ථි (Susman 2008) සොයා ගැනීම, මෙම මානව ෆොසිල පිලිබද ව කතුවරුන් තුළ වූ කුතුහලය වැඩි කිරීමට පෙලඹී ඇත. මේ කාලය වන තුරු, Homo erectus ට වඩා ප්රාථමික ලක්ෂණ පෙන්නුම් කළ හෝමෝ නිදර්ශක වාර්තා වී නොමැත. පසුකාලීන ව, නැගෙනහිර අප්රිකාවේ මුල් හෝමෝ ගණයට අදාල සොයාගැනීම් හි අතොරක් නැති විය (Leakey 1961, 1971, 1973a; b; Day 1971; Day & Leakey 1973; Day et al. 1976; Johanson et al. 1987). මෙම නව ෆොසිල හැබිලිස් මානවයා පිළිබද වූ පුළුල් විද්යාත්මක පිළිගැනීමට දායක වූ නමුත්, එය වසර 15ක ප්රතිරෝධයකින් පසුව ස්ථාපිත වීම විශේෂ කරුණකි (Tobias 2009).
හෝමෝ කුලයට අයත් ආදිතම සාමාජිකයන් අතරින් වඩාත් ප්රසිද්ධ විශේෂ වන්නේ Homo habilis, Homo rudolfensis, හා Homo erectus (වගුව 1) ය. හෝමෝ ඉරෙක්ටස් හා සසදන විට හෝමෝ හැබිලිස් හා හෝමෝ රුඩොල්ෆෙන්සිස් යන විශේෂ වඩාත් ප්රාථමික මට්ටමේ පරිණාමීය ලක්ෂණ දරයි. බොහෝ විට ඔවුන් එකිනෙකාගෙන් ව්යුහ විද්යාව සහ හැසිරීම් වලින් වෙනස් වන ආකාරය සහ පෙර සිටි ඔස්ට්රලෝපිතකස්වරුන්ගෙන් වෙනස් වන්නේ කෙසේද යන්න පිලිබද ව පර්යේෂණ සිදුවී ඇත්තේ අල්ප වශයෙනි. නමුත් අවබෝධ කරගතයුතු කරුණ වන්නේ නූතන මිනිසුන් හා සසඳන විට එය ව්යුහ විද්යාත්මකව සහ චර්යාත්මකව (anatomically and behaviorally) ප්රාථමික වුවද, මෙම මානව විශේෂ තුන, මානව පරිණාමීය ගමනේ, විශේෂයෙන් මොළයේ සහ ශරීරයේ ප්රමාණය වැඩි වීම සහ වඩ වඩාත් සංකීර්ණ මෙවලම් භාවිතය හා හැසිරවීම හරහා විශාල ඉදිරි පිම්මක් සනිටුහන් කරන බවය.
වගුව 1- හොමෝ හැබිලිස්, හෝමෝ රුඩොල්ෆෙන්සිස් හා හෝමෝ ඉරෙක්ටස් මානවයින්ගේ තොරතුරු
| මානව විශේෂය | වසර මිලියන | ප්රධාන ෆොසිල | පූර්ව නාමකරණ | ප්රධාන භූගෝලීය කලාප |
| හෝමෝ හැබිලිස්
Homo habilis |
2.4–1.4 | KNM-ER 1805 (cranium); KNM-ER 1813 (cranium); KNM-ER 42703 (maxilla); OH 7 (type; parts of skull, hand); OH 8 (foot); OH 16 (cranium); OH 24 (cranium); OH 62 (partial skeleton); OH 65 (maxilla); SK 847 (cranium) | Australopithecus habilis; Homo rudolfensis | Omo, Ethiopia; Ileret and Koobi Fora, Kenya; Olduvai Gorge, Tanzania; Drimolen, Sterkfontein, and Swartkrans, South Africa |
| හෝමෝ රුඩොල්ෆෙන්සිස්
Homo rudolfensis |
1.9 | UR 501 (mandible) | Homo habilis; Australopithecus rudolfensis | Koobi Fora, Kenya; Uraha, Malawi |
| හෝමෝ ඉරෙක්ටස්
Homo erectus |
1.8–0.4 | Perning 1 (calvaria); Ngandong 7 (cranial vault); Ngandong 13 (cranial vault); Sangiran 2 (cranial vault); Sangiran 17 (cranium); Trinil 2 (type; skullcap); Zhoukoudian XII (calvaria); D2280 (calvaria); D2282 (cranium); D2700 (cranium); D3444 (cranium); KNM-ER 992 (mandible); KNM-ER 1481 (femur); KNM-ER 3733 (cranium); KNM-ER 3883 (cranium); KNM-ER 42700 (cranial vault); KNM-OG 45500 (cranial vault); KNM-WT 15000 (skeleton); OH 9 (cranial vault); OH 28 (partial pelvis) | Homo ergaster; Homo georgicus; Homo erectus sensu stricto | Mojokerto, Ngandong, Sambungmachan, Sangiran, and Trinil, Java, Indonesia; Zhoukoudian, Hexian, Nanjing, and Gongwangling, China; Narmada, India; Dmanisi, Georgia; Salé, Morocco; Buia, Eritrea; Daka, Middle Awash, Ethiopia; Ileret, Koobi Fora, Olorgesailie, and West Turkana, Kenya; Olduvai Gorge, Tanzania; Sterkfontein and Swartkrans, South Africa
|
වසර මිලියන 2.3 සිට 1.4 දක්වා කාලයට අයත් වන හෝමෝ හැබිලිස් මානවයා, ඔස්ට්රලෝපිතිකස් විශේෂයකින් පැවත එන බව සාමාන්යයෙන් දන්නා මතය වේ. හෝමෝ හැබිලිස් නැඟෙනහිර අප්රිකාවේ පරිණාමය වූයේ නම්, ඔස්ට්රලෝපිතිකස් ඇෆරෙන්සිස් සහ ඔස්ට්රලෝපිතිකස් ගාහි යන පූර්වජයන් ගුරුකොට පරිණාමය වී ඇත, අනෙක් අතට හෝමෝ හැබිලිස් දකුණු අප්රිකාවේ පරිණාමය වූයේ නම්, ඔස්ට්රලෝපිතිකස් ඇෆිරිකානස් හා ඔස්ට්රලෝපිතිකස් සෙඩීබා යන මානවයින් පූර්වජ අපේක්ෂකයන් ලෙස හැදින්විය හැකි ය. මෙම ගැටළුවට පිළිතුරු අත්යවශයෙන් ම අනාගත පර්යේෂණ මත රදා පවතිනු ඇත. කෙසේ වෙතත්, හෝමෝ හැබිලිස් මානවයා ගැන දැනට සිදුවී ඇති පර්යේෂණ මත පදනම්ව, ඔහු හෝමෝ සේපියන් ඇතුළු හෝමෝ කුලයේ පසුකාලීන සියලුම විශේෂවල පොදු පූර්වජයෙකු බව බොහෝ දෙනා පිළිගෙන ඇත.

Leakey, Tobias සහ Napier දිගින් දිගටම තර්ක කළේ ඔවුන් සොයා ගත් ෆොසිල, හෝමෝ ඉරෙක්ටස් තරම් විශාල මොළයක් හෝ ශරීරයක් නොමැති බවත්, එය ඔස්ට්රලෝපිතකස් තරම් ප්රාථමික හා වානර නොවන බවයි. මේ අනුව, ඔවුන් හෝමෝ ගණයට අයත් නිදර්ශක සඳහා මූලික අවශ්යතා කිහිපයක් හදුන්වා දෙන ලදී. පළමුවෙන්ම, මොළයේ ප්රමාණය ඝන සෙන්ටිමීටර 600 ට වඩා වැඩි විය යුතු අතර, එය සියලුම ඔස්ට්රලෝපිතිකස් නිදර්ශකවල කපාල ධාරිතාවයට වඩා විශාලය. ඊට අමතරව, හිස් කබලේ අස්ථි සුමට හා වටකුරු විය යුතු අතර, ඔස්ට්රලෝපිතිකස් ප්රදර්ශනය කරන ලද ලෝම අවම හෝ නොමැති විය යුතු ය. හිස් කබල රූප විද්යාවෙන් (cranial morphology) ඔබ්බට, හෝමෝ ගණයට අයත් වීමේ දෙවන නිර්ණායකය ලෙස ඔවුන් තර්ක කළේ, විශාල මොළය හා සම්බන්ධ චර්යාත්මක සහ සංජානන සංකීර්ණත්වයේ වැඩි වීමක් පෙන්නුම් කරන ශිලාමෙවලම් සෑදීමේ හැකියාව ඔවුන් සතු විය යුතු බවයි. ශිලාමෙවලම් සහිත සන්දර්භයකින් හෝමෝ ෆොසිල සැකිල්ලක් කිසිවෙකු වර්තමානය වන විටත් සොයාගෙන නැතත්, අසල ඇති ශිලාමෙවලම්වල නිර්මාතෘන් ඔවුන් යැයි පුරාවිද්යාඥයින් නිගමනය කරයි.
තවදුරටත් Leakey සහ සගයන් යෝජනා කළේ හෝමෝ ගණයේ මුහුණ සහ හකු ප්රදේශය, ඔස්ට්රලෝපිතකස් විශේෂයට වලට වඩා කුඩා විය යුතු බවත්, සහ පශ්චාත් කපාල (postcranial) සැකිල්ල නූතන සේපියන් මානවයින්ට සමාන විය යුතු බවත් ය. එමෙන් ම හෝමෝ හැබිලිස් මානවයාගේ නිත්ය වූ ඍජු චලනයක් තිබියදීත්, තවමත් ප්රාථමික ගති ලක්ෂණ (තරමක් අපසරනය වන මහපටැඟිල්ලක් වැනි ලක්ෂණ) තිබූ අතර, පෙර ඔස්ට්රලෝපිතිකස් සාමාජිකයින් මෙන් නොව, දෙපයින් ඇවිදීම සඳහා පාද සම්පූර්ණයෙන්ම අනුවර්තනය වී ඇති බව පැහැදිලි වේ. හෝමෝ ගණයේ ෆොසිල වාර්තාෙවේ වඩාත් කැපී පෙනෙන ලක්ෂණ ලෙස සම්පූර්ණ ලෙස ද්විපාදකරණයට ලක් වීම හා විශාල කපාල ධාරිතාවක් සහිත මොළයක් තිබීම දැක්විය හැකි ය. මෙම ගති ලක්ෂණ සමූහයට අමතර ව, සමස්තයක් වශයෙන් කුඩා දත් පිහිටීම, (කෘන්තක හා සසඳන විට සාපේක්ෂ කුඩා චාර්වක සහ පුරස්චාර්වක දත් පිහිටීම), තවදුරටත් ප්රමාණයෙන් කුඩා වූ රදනක දත්, ඝන එනැමලය, හා පරාවලයික දන්ත වක්ර (parabolic dental arch) පිහිටයි. එනම්, දත් කලින් මානව සහ මානව නොවන වානරයන්ගේ මෙන් v-හැඩය හෝ u-හැඩය වෙනුවට අශ්වලාඩමක (horseshoe shape) හැඩයක් සාදයි.

හෝමෝ හැබිලිස් මානවයා
හෝමෝ හැබිලිස් යනු කුඩා ප්රමාණයේ සිරුරක් සහිත මානවයෙක් වූ අතර, ඔස්ට්රලෝපිතෙකස් සහ පරන්ත්රොපස් විශේෂවලට වලට වඩා විශාල මොළයක් (ඝන සෙන්ටිමීටර 600කට වඩා වැඩි) සහිත විය. කෙසේ වෙතත්, KNM-ER 1813 වැනි නිදර්ශක ආශ්රිත හිස්කබලේ ධාරිතාව ඝන සෙන්ටිමීටර 510ක් පමණක් නිසා, විචල්යමය පරාසය පිළිබද යම් යම් වෙනස්කම් දැකිය හැකි ය (Miller 1991). එම නිදර්ශකය සතු මොලයේ වටකුරු බව හා චාර්වක දත් පටු වීම හා තෙවන චාර්වක දත ප්රමාණයෙන් කුඩා වීම මනුෂ්යත්වය ලගාකරගැනීම පිළිබද මුල්කාලීන ඉගි සපයයි (Wood 1992). ඔවුන්ගේ දන්ත එනමලය තවමත් ඝන වූ අතර හකු තවමත් ශක්තිමත් වීම තුළින් පෙන්නුම් කරන්නේ ඔවුන් දෘඩ ආහාර වර්ග කිහිපයක් වෙත අනුවර්තනය වෙමින් පැවති බවයි (Haeusler & McHenry 2004). ක්ෂුද්ර දන්ත අධ්යයන (Dental microwear) අධ්යයනවලින් පෙනී යන්නේ හෝමෝ හැබිලිස් මානවයාගේ ආහාර නම්යශීලී වූ අතර ඔවුන් කොළ, දැවමය ශාක සහ සමහර සත්ව පටක වැනි දැඩි ආහාර ඇතුළු පුළුල් පරාසයක ආහාර අනුභව කිරීමට සමත් වූ නමුත් ඔවුන් ඒවා නිතිපතා පරිභෝජනය නොකළ බවයි (Ungar et al. 2006).


රූපය 3 – හොමෝ හැබිලිස් මානව ෆොසිලයක් හා ජාන විද්යාත්මක සාධක අනුව සිදුකරන ලද ප්රතිනිර්මිත හෝමෝ හැබිලිස් මානවයා
හෝමෝ හැබිලිස් මානවයාගේ ආහාර රටාව සඳහා තවත් සාක්ෂියක් ලැබෙන්නේ මීට වසර මිලියන 2.6කට පෙර සොයාගත් පැරණිතම කැපුම් සහ සීරුම් ලකුණු සහිත අස්ථි කිහිපයකිනි. විශාල සතුන්ගේ මාංශ හා අස්ථි වෙන් කිරීමේ සලකුණු සමග මස් සහ ඇට මිදුළු අනුභව කිරීම පිළිබඳ සෘජු සාක්ෂි හෝමෝ හැබිලිස් මානවයාගේ ෆොසිල සාක්ෂි සමග සමකාලීන වේ (Domínguez-Rodrigo et al. 2005). බොහෝ විද්යාඥයන්ගේ මතය වන්නේ හැබිලිස් ඇතුළු මුල් හෝමෝවරුන් විසින් පුරාවිද්යා වාර්තාවල ඇති මුල්ම ශිලාමෙවලම් සාදා භාවිත කළ බවයි. කෙසේ වෙතත්, මෙම උපකල්පනය පරීක්ෂා කිරීමට අපහසු වන්නේ මුල් මානව වර්ග කිහිපයක් එකම කාලයකදී සහ එකම භූගෝලීය ප්රදේශයක, පැරණිතම මෙවලම් භාවිතය ආශ්රිත සලකුණු සොයාගෙන ඇති බැවිනි (Bobe 2004; Domínguez-Rodrigo et al. 2005).
ඔල්දුවායි ගෝජ් (Olduvai Gorge) හි OH 62 දරන අර්ධ ලෙස සංරක්ෂණය වූ නිදර්ශකය (Johanson et al. 1987) ඇසුරෙන් සිදුකර ඇති විද්යාත්මක නිගමන අනුව එම හෝමෝ හැබිලිස් නිදර්ශකයේ ශරීර ප්රමාණය සහ අත් පා සමානුපාත ය, හෝමෝ ඉරෙක්ටස් ගේ නම් එම අනුපාතය විෂම වේ (Haeusler & McHenry 2004; Larson 2009). මෙම විශේෂය සඳහා පශ්චාත් හිස්කබල් ෆොසිල (post cranial fossils) හිඟකම සැලකිල්ලට ගෙන ශරීර ස්කන්ධය ඇස්තමේන්තු කිරීම අපහසු ය, සාමාන්යයෙන් පිළිගත් ශරීර බර ඇස්තමේන්තුව කිලෝග්රෑම් 34ක් වන අතර, එම අගය ඔස්ට්රලෝපිතෙකස් සහ පරන්ත්රොපස් සඳහා වන ඇස්තමේන්තුගත ස්කන්ධයට වඩා කුඩාය (කිලෝග්රෑම් 44) සහ හොමෝ ඉරෙක්ටස් (කිලෝග්රෑම් 56- 75) ට වඩා කුඩා වේ (Ruff et al. 1997).
හෝමෝ රුඩොල්ෆෙන්සිස් මානවයා (Homo rudolfensis)
හෝමෝ ඉරෙකටස් හා හෝමෝ හැබිල්ස් යන මානව විශේෂවල අතරමැදි ලක්ෂණ දරන හෝමෝ රුඩොල්ෆෙන්සිස් මානවයා පිලිබද අනාවරණය වී ඇත්තේ අල්ප වශයෙනි. මෙම විශේෂය නියෝජනය කරනු ලබන්නේ කෙන්යාවේ කූබිෆෝරා (Koobi Fora) (KNM-ER 1470) හි දත් රහිත හිස් කබලකින් සහ මලාවි (Malawi) (UR 501) හි හකු අස්ථියකින් පමණි. KNM-ER 1470 නිදර්ශක අංකය දරන හිස්කබලේ ධාරිතාව ඝන සෙන්ටිමීටර 752ක් (Holloway 1983) ලෙස ඇස්තමේන්තු කර ඇති අතර, දත් දුර්වල ලෙස සංරක්ෂණය වී ඇති නමුත්, දත් කුහර (alveoli or tooth sockets) විශාල වන අතර, ඒවායේ අඩංගු වූ දත්වල ප්රමාණයද එමගින් නිර්ණය කරගත හැකි ය (Wood & Collard 1999).
හෝමෝ හැබිලිස් හා සසඳන විට හෝමෝ රුඩොල්ෆෙන්සිස් මානවයා හට විශාල සහ පළල් වූ දත් තිබී ඇත. ඔවුන්ගේ දත් ඔස්ට්රලෝපිතිකස් සාමාජිකයින් අතර දක්නට ලැබෙන ඒවාට වඩා මදක් කුඩා වූ අතර, හෝමෝ රුඩොල්ෆෙන්සිස් මානවයා හට මුල් මිනිසුන් තුළ දක්නට ලැබෙන දැඩි ලෙස ගොඩනඟන ලද හකු සහ ශක්තිමත් හකු මාංශ පේශී බැඳීම් නොතිබුණි. මෙම ව්යුහ විද්යාත්මක වෙනස්කම් හෝමෝ රුඩොල්ෆෙන්සිස් සහ පෙර පූර්වජයින් අතර වූ ආහාර විවිධත්වය පෙන්නුම් කරයි. අනෙකුත් මුල් හෝමෝ විශේෂ මෙන්ම හෝමෝ රුඩොල්ෆෙන්සිස් මානවයා ද ඔවුන්ගේ ආහාර සැකසීමට ශිලාමෙවලම් භාවිතා කරන්නට ඇත. කෙසේ වෙතත්, මෙවලම් නිෂ්පාදනය සහ භාවිතය ආරම්භ වූ කාලයේ මුල් මානව වර්ග එකකට වඩා වැඩි ගණනක් ජීවත් වූ බැවින්, පළමු ශිලා මෙවලම් සෑදීමට සහ භාවිතා කිරීමට වගකිව යුත්තේ කුමන විශේෂද යන්න විද්යාඥයින්ට නිශ්චිතව පැවසීමට අපහසුය. හෝමෝ රුඩොල්ෆෙන්සිස් මානව ෆොසිල හා සමාන්තර ස්ථරවල දැනට ශිලා මෙවලම් හමු වී නැත, නමුත් හෝමෝ රුඩොල්ෆෙන්සිස් මානවයා ජීවත් වූ එම කාල වකවානුවේම පවතින ශිලා මෙවලම් අනාවරණය වී ඇත.
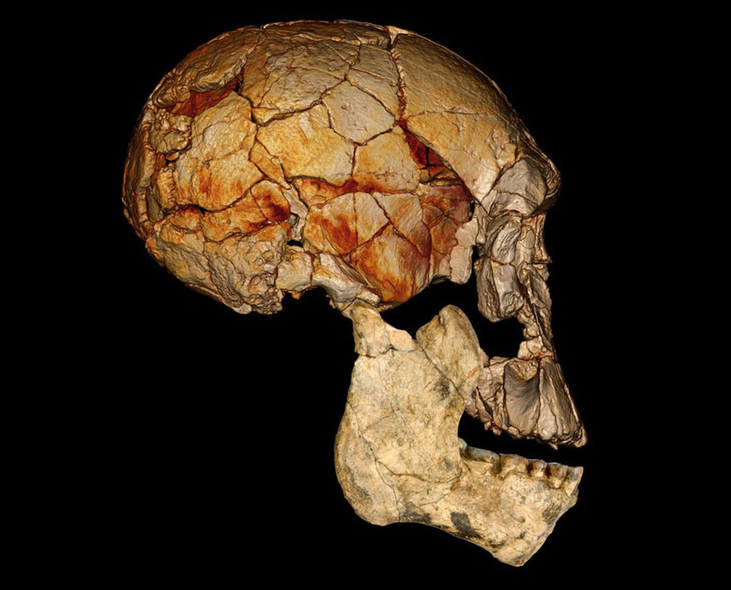
හෝමෝ ඉරෙක්ටස් මානවයා (Homo erectus)
ලන්දේසි ජාතික Eugene Dubois ඉන්දුනීසියාවේ ජාවා දූපතේ Trinil නදි නිම්නයෙන් හෝමෝ ඉරෙක්ටස් මානවයාගේ පළමු ෆොසිල සොයා ගන්නා ලදී (Baur 1895). මුල්ම නාමකරණයේ දී Pithecanthropus erectus ලෙස හැඳින්වූ අතර, හිස් කබලේ ඉහළ කොටස (top of the brain case and the browridges) කුඩා හා ප්රාථමික පෙනුමක් ඇති, විශේෂයෙන් එය පෙර මානව පූර්වජයින් සමග සංසන්දනය කළ නොහැකි තරම් බාහිර රූපවිද්යාත්මකව වෙනස් ලක්ෂණ පෙන්නුම් කෙරිණි (Dunsworth 2010). කෙසේ වෙතත්, හිස්කබලේ වූ බාහිර රූපවිද්යාත්මක ලක්ෂණ හා ගාත්රා අස්ථි පැහැදිලිවම සෘජු, විශාල ශරීරයක් ඇති මානවයතුට අයත් බව හැගවිණි. මෙකී මූලික සොයාගැනීම්වලට අමතරව තවත් ඉරෙක්ටස් ෆොසිල ඉන්දුනීසියාව (Sartono 1972), චීනය (Black 1927, 1934; Weidenreich 1944; Clarke et al. 1970; Hughes & Tobias 1977), උතුරු අප්රිකාව (Arambourg 1955; Arambourg & Biberson 1956), ජෝජියාව (Gabunia & Vekua 1995; Gabunia et al. 2000; Lordkipanidze et al. 2005, 2007; Rightmire et al. 2006) හා ඉන්දියාව (Sonakia & Kennedy 1985) ආශ්රයෙන් ද සොයා ගැනිණි. කෙසේ වෙතත්, හෝමෝ ඉරෙක්ටස් මානවයෙක් සතු වූ හොඳම තනි නිදර්ශකය වන්නේ කෙන්යාවේ බටහිර ටර්කානා නිම්නයේ Nariokotome ක්ෂේත්රයෙන් හමු වූ පිරිමි ළමයෙකුට අයත් සම්පූර්ණ ෆොසිල සැකිල්ලයි (Brown et al. 1985).

විශාල ශරීර ප්රමාණය, විශාල මොළය සහ කුඩා දත් සහිත, බොහෝ හෝමෝ ඉරෙක්ටස් මානව ෆොසිල අනෙකුත් මුල් හෝමෝ විශේෂ දෙකෙන් වෙනස් වේ. හෝමෝ ඉරෙක්ටස් මානවයාට අයත් දත් හෝමෝ හැබිලිස් සහ හෝමෝ රුඩොල්ෆෙන්සිස් මානව විශේෂවලට වඩා කුඩා වන අතර මුල් නිදර්ශක වල විශාල දත් තිබුණද, කාලයත් සමඟ දතේ ප්රමාණය අඩු වීමක් නිරීක්ෂණය කළ හැකි ය. හෝමෝ ඉරෙක්ටස් මානවයාට බොහෝ නිදර්ශකවල ඉහළ කෘන්තක දත සවලක් ආකාරයේ පැතලි හැඩැති (shovel) වන අතර නූතන මිනිසුන් තුළ ද දක්නට ලැබෙන ලක්ෂණයක් ලෙස ගිණිය හැකි ය.
හෝමෝ ඉරෙක්ටස් මානවයාගේ කපාල ධාරිතාවය ඝන කියුබික් සෙන්ටිමීටර් 700 – 1300ත් අතර අගයක් ගන්නා අතර එය පැහැදිලිවම පූර්වජයන්ට වඩා වෙනස් ආකාරයක් ගනී. බාහිර රූපවිද්යාත්මක ව වුවද හිස්කබලේ ඝන ඇහිබැමක් නිරික්ෂණය කළ හැකි අතර, අන් මානවයන්ගෙන් වෙනස් වූ supra-orbital torus නම් අස්ථි ප්රදේශයකට ප්රවේශ වේ. එමෙන් ම නළල ප්රදේශය තරමක් පසුපසට විහිදුණු ආකාරයක් ද වේ. මූලිකවම හිස්කබල දිගින් වැඩි හා උසින් අඩු (long and low) ස්වභාවයක් ගන්නා අතර නූතන මානවයාගේ එය මුළුමනින්ම රවුම් ආකාර ගනී. භාෂාවක් හෝ සන්නිවේදන කටයුතු භාවිත කරන ලද පළමු හෝමෝ සාමාජිකයා ලෙස ඉරෙක්ටස් හැදින්වෙන්නේ ඔවුන්ගේ ෆොසිල කායච්ඡේදවිදයාත්මකව අධ්යයනයේ දී හමු වූ Hyoid අස්ථිය නිසාවෙනි. එනම් එම අස්ථිය මගින් හඩක් නිපදවීමේ හැකියාව ඉරෙක්ටස් මානවයාට තිබූ බව සනාථ වේ (Capasso et al. 2008). හෝමෝ ඉරෙක්ටස් මානවයා ආහාර දාමයේ මුල් පුරුකක් ලෙස ක්රියාකරන ලද අග්ර විලෝපිකයෙකි. ඒ බව ඔහු දඩයම් කරන ලද විශාල හා මධ්යම ප්රමාණයේ සතුන් (අලි ඇතුන්, බොවිඩේ කුලයට අයත් සතුන්) තුළින් මූර්තිමත් වේ.
හෝමෝ ඉරෙක්ටස් මානවයාගේ පරිණාමය සමග Acheulean ශිලාමෙවලම් කර්මාන්තය අතිශයින් බද්ධ වී පවතින අතර, ගින්දර භාවිතා කිරීමට, දඩයම් කිරීමට සහ කණ්ඩායම් ලෙස එක්රැස් වීමට, තුවාල වූ හෝ අසනීප වූ කණ්ඩායම් සාමාජිකයින්ට රැකබලා ගැනීමට සහ සමහර විට මුහුදු ගමන් සිදුකිරීමට සහ කලාව කෙරෙහි වූ හැකියාව අත්කරගත් මුල්ම මානව පූර්වජයින් ලෙස හෝමෝ ඉරෙක්ටස් මානවයා හැදින්වේ.
මුල් හෝමෝවරුන්ගේ චර්යාත්මක ලක්ෂණ
මුල් හෝමෝවරුන්ගේ හැසිරීම් හි සංකීර්ණත්වය සඳහා උදාහරණ ලැබෙන්නේ මානව ෆොසිල හා සම්බන්ධ පුරාවිද්යාත්මක අවශේෂ මගිනි. ලතින් නාමයෙන් පරිවර්තනය කර ඇති පරිදි, හෝමෝ හැබිලිස් හෙවත් Handy man, පළමු ශිලා මෙවලම් නිෂ්පාදකයා ලෙස සැලකේ. ඔවුන් ප්රාථමික ශිලාමෙවලම් නිර්මාණය කරන ලද අතර, Olduvai Gorge ආශ්රිතව සොයාගැනුණු මූලික සොයාගැනීම් තුලින් එම තාක්ෂණය ඔල්දුවාන් කර්මාන්තය (Oldowan industry) ලෙස නම් කරන ලදී. මාංශ වෙන් කර ගැනීම සඳහා භාවිත කරන ලද කෙටීම් ආයුධ, පතුරු ආයුධ, න්යාෂ්ටි සහ මිටි ගල් වැනි ආයුධ, සත්ත්වපුරාවිද්යාත්මක සාක්ෂි සමඟ සම්බන්ධ වන්නේ කලාතුරකිනි (Roche et al. 2009). එපමණක් නොව, මුල් හෝමෝ බොහෝ විට ගොදුරු දඩයම් කිරීමට වඩා සත්ව මළකුණු මත යැපුණු බව අනාවරණය වේ. පසුකාලීනව ඇචුලියන් මෙවලම් භූගෝලීය වශයෙන් පුළුල්ව භාවිතා කිරීමෙන් ඇඟවෙන්නේ ඒවා මාංශ සකස් කිරීමට පමණක් සීමා නොවී බහුකාර්යය මෙවලම් කට්ටලයක් ලෙස භාවිතයේ පැවති බව ය (Roche et al. 2009). උදාහරණයක් ලෙස, ශාක ද්රව්ය සැකසීම සඳහා ද ශිලා මෙවලම් මහෝපකාරී වනු ඇත. Oldowan ආයුධ, ප්රාථමික හා සරල වුවද, කාලය සහ පතුරු නිෂ්පාදනයේ දී නිරීක්ෂණය වූ විචල්යතා හරහා වෙනස් වූ බව පෙන්නුම් කෙරේ (Braun et al. 2008a; b; Roche et al. 2009).

මානව පරිණාමීය ගමන් මගේ මුල්ම ශිලා මෙවලම් භාවිතය අදින් වසර මිලියන 2.6ක් පැරණි අවසාදිත ඇසුරේ සොයාගන්නා ලදී (Semaw et al. 2003). සත්ත්ව අස්ථි මත වූ කැපුම් ලකුණු ඔස්ට්රලෝපිතකස් ගාහි ට අයත් ෆොසිල අවශේෂ ඇසුරේ අර්ථකතනය කළ අතර (Heinzelin et al. 1999), එය එසේ වී නම් පළමු හෝමෝ ගණයේ සාමාජිකයාගේ පහළ වීම තවත් වසර ලක්ෂයක් පමණ ආපස්සට ගැනීමේ හැකියාව ඇත. නමුත් ඒ සදහා ප්රමාණවත් පුරාවිද්යාත්මක සාක්ෂි මෙතෙක් ගොනු වී නැත. තවදුරටත් පලමු ශිලා මෙවලම් නිෂ්පාදනය කිරීමේ ගෞරවය හිමිවන්නේ හෝමෝ හැබිලිස් හට ය.
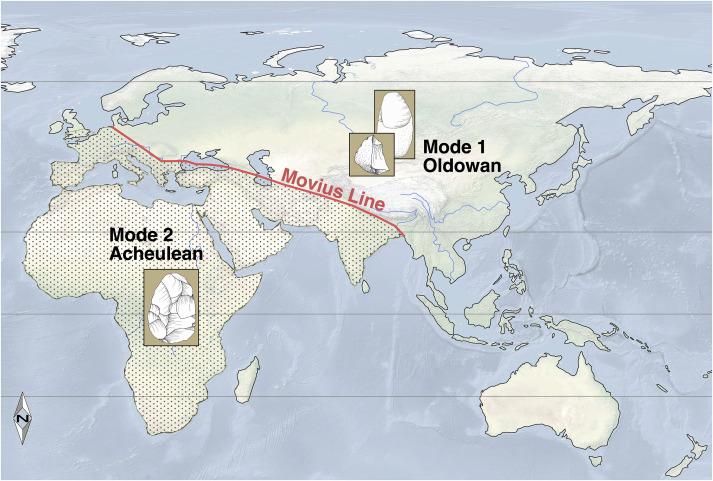
ශිලා මෙවලම් වාර්තාවේ මෙම මීළඟ තාක්ෂණික අදියර ඇචුලියන් ආයුධ වන අතර හෝමෝ ඉරෙක්ටස් හා අප්රිකානු මහද්වීපය සමඟ සමීපව සම්බන්ධ වේ. මෙම නව තාක්ෂණයේ විශේෂ අංගය ලෙස කඳුළු බිංදුවක හැඩැති ද්විමුහුණත් “අත්පොරොව” හැඳින්වේ (රූපය 5). ඒවා තියුණු දාරවලින් සමන්විත බැවින්, ශාක ද්රව්ය සැකසීමට සහ සතුන්ගේ සම, මාංශ හා අස්ථි එකිනෙකින් වෙන්කර ගැනීම සඳහා අත්පොරොව භාවිතා කර ඇති බවට සාක්ෂි හමුවේ. වඩාත් දියුණු ඇචුලියන් මෙවලම් – ඒවා මුලින් සොයාගත් ප්රංශයේ ශාන්ත ඇචූලියන් පුරාවිද්යා ක්ෂේත්රය සමග නම් කර ඇත. Oldowan මෙවලම් වලට වඩා නිර්මාණ කාර්යය දුෂ්කර වූ ඇචුලියන් මෙවලම් පසුපස හෝමෝ ඉරෙක්ටස් මානවයා අත් කරගත් සංජානන හා චර්යාත්මක සංකීර්ණත්වය නිරූපණය වේ. හෝමෝ ඉරෙක්ටස් මානව ෆොසිල හමුවන සෑම භුගෝලීය ප්රදේශයකින්ම ඇඩුලියන් ආයුධ හමු නොවේ, උදාහරණ ලෙස ජෝජියාවේ Dmanisi හා නැගෙනහිර ඉන්දියාවෙන් බහුලව හෝමෝ ඉරෙක්ටස් ෆොසිල හමු වුවද ඇචුලියන් තාක්ෂණය අදාල ද්රව්යාත්මක සාක්ෂි හමුවන්නේ අල්ප වශයෙනි. ඒ සදහා විය හැකි උපකල්පනයක් ලෙස පුරාමානව විද්යාඥයින් පෙන්වා දෙන්නේ ඉරෙක්ටස් මානවයා නැගෙනහිර හා අග්නිදිග ආසියාවට විසිරී යනන්ට පෙර ඔවුන් වෙනත් අමුද්රව්ය භාවිත කරමින් ආයුධ නිපදවන්නට ඇති බවත්, එම ප්රදේශවල ඇති නිවර්තන දේශගුණය හේතුවෙන් අද වන විට ඒවා පුරාවිද්යාත්මක වාර්තාව තුල දක්නට නොවන බවත් ය. දැනට මෙම පෙරදිග-බටහිර සංසිද්ධිය පිළිබඳ විවාදය දිගටම පවතින අතර, එය මුලින්ම රටාව නිරීක්ෂණය කළ පුරාවිද්යාඥයාගේ නමින් Movius Line ලෙස හදුන්වනු ලබන මනංකල්පිත රේඛාව ඇසුරෙන් ඉරෙක්ටස් ව්යාප්තිය බෙදා දක්වනු ලබයි (Movius 1948).

ශිලා මෙවලම් තාක්ෂණයේ දියුණුවත් සමඟ, හෝමෝ ඉරෙක්ටස් මානවයා මාංශ භාවිතය වැඩි කළ බවට කායච්ඡේදවිද්යාත්මක සාක්ෂි ඇත. හෝමෝ ඉරෙක්ටස් මානවයාගේ චාර්වක දත් මත ඇති තුණ්ඩ (cusps), ඉන්පෙර සිටි මානව ජූර්වජයින්ගේ චාර්වක දත්වල ඇති රටාවට වඩා දැඩි වන අතර, මෙම දන්ත විෂමතාවය මාංශ කැබලි කරගැනීමට (Ungar 2004, 2012) හෝ පොදුවේ දැඩි ආහාර කැඩීමට ප්රයෝජනවත් වනු ඇත (Ungar & Scott 2009). කෙසේ වෙතත්, ක්ෂුද්ර දන්තවිද්යාත්මක (dental microwear) අධ්යයනවලින් ලැබෙන සාක්ෂි, මුල් හෝමෝවරු නිරන්තරයෙන් දැඩි ආහාර අනුභව කළ බවට පූර්ණ සහාය නොදක්වයි. කෙසේ වෙතත්, පොදුවේ ගත් කල, මෙම මානවයින් ජීවත් වූ විවිධ පරිසර තත්ත්වවලට අනුව දන්ත රූපවිද්යාත්මක සාක්ෂි විචලනය වූ බව නිගමනය කළ හැකි ය (Ungar & Scott 2009).
පාරිසරික වෙනස් වීම් හා මුල්ම හෝමෝවරුන්
Plio-Pleistocene කාලපරිච්ඡේදයේ සංක්රාන්තිය ගෝලීය දේශගුණික විපර්යාස හේතුවෙන් නැගෙනහිර අප්රිකානු පරිසරයේ විශාල වෙනසක් සනිටුහන් කරයි (deMenocal 2004; Maslin & Trauth 2009). මෙම කාල පරිච්ඡේදය තුළ දේශගුණික විචල්යතාවය, ක්ෂීරපායී ෆොසිල වාර්තා, Plio-Pleistocene මානව පරිණාමය, විචලනය සහ අනුවර්තනය සමඟ සම්බන්ධ වී ඇතැයි උපකල්පනය කෙරේ (Potts 1998a; b; Maslin & Trauth 2009). සිසිල් වූ Plio-Pleistocene දේශගුණය අප්රිකානු වැසි වනාන්තර, උප සහරා අප්රිකාවේ බහුලව පැතිරී ඇති සැවානා තණබිම් වලට ව්යාප්ත වීමට ඉඩ හැරියේ ය. මෙම මුල් සැවානා විශාල තෘණ ශාක භක්ෂක සතුන්ගෙන් ගහණ වූ අතර එය හෝමෝ ඉරෙක්ටස් මානවයා සඳහාම වූ නිත්ය ආහාර සංචිතයක් බදු විය – බොහෝ විට මෙම වනාන්තර නොවන පරිසර කෙරෙහි (සැවාණා තෘණ භූමි) දැඩි ලෙස අනුවර්තනය වූ පළමු මානව වර්ගය ලෙස ද ඉරෙක්ටස් මානවයා හැදින්වේ (Reed 1997). වසර මිලියන 2.5 ත් 2 ත් අතර කාලයට පෙර ශුෂ්ක, සෘතුමය තෙත් සහ වියලි කලාපීය වාසස්ථාන පැරණිතම හෝමෝවරුන් විසින් පරිභෝජනය කළ අතර, ඉන් පසුව මානවයින් වැඩි වැඩියෙන් විවෘත හා තෙත් තණබිම් වලට වැඩි නැඹුරුවක් දැක්වීය (Reed & Russak 2009). Koobi Fora හි (Braun et al. 2010) හි වසර මිලියන 1.9 ක් පැරණි ක්ෂේත්රයකදී අනුභව කරන ලද කැස්බෑවන්, කිඹුලන් සහ මත්ස්යයින් මෙම පුරාණ පාරිසරික අර්ථ නිරූපණය හා සාධාරණීකරණයට සහාය දක්වයි. හෝමෝ ඉරෙක්ටස් මානවයාගේ දන්ත එනමලයේ ජෛව භූ රසායනික විශ්ලේෂණය තුළින් (Biogeochemical analysis of tooth enamel) හෝමෝ ඉරෙක්ටස් මානවයාගේ බාහිර පෙනුම සහ ව්යාප්තිය වැඩිවන තෘණ බිම් පරිසරයකට සම්බන්ධ කරයි(Bobe & Leakey 2009; Sponheimer & Lee-Thorp 2009).
තව දුරටත් සත්ත්වපුරාවිද්යාත්මක සාක්ෂි මගින් හෝමෝ ඉරෙක්ටස් මානවයාගේ ආහාර රටාව තුළ සත්ව ප්රෝටීන් වැඩි වශයෙන් අඩංගු බව සනාථ කර ඇත. විශාල ක්ෂීරපායී අස්ථිවල ශිලා මෙවලම්වලින් කැපූ බවට සලකුණු නිරීක්ෂණය කළ හැකි අතර, එලෙස අස්ථි ආශ්රිත පුරාවිද්යාත්මක සන්දර්භ ඇසුරේ හෝමෝ ඉරෙක්ටස් මානවයාට අදාල පුරාකෘති පැතිරීම බහුල වේ (Roche et al. 2009). ආදිතම මානවයින් මාංශ අනුභව කිරීමේ සහ ආහාර පිසීමේ හැකියාව පිළිබඳ සාක්ෂි අතර දිගු පරතරයක් පවතී. තවද සහ ඉහත සඳහන් කළ පරිදි, හෝමෝ ඉරෙක්ටස් මානවයාට හට සිය දඩයම් උපාය මාර්ගයට ප්රක්ෂේපන (projectiles) සාර්ථකව ඇතුළත් කිරීමේ හැකියාව නොතිබෙන්නට ඇත. ශරීරය ද්විපාදකරණය සඳහා අනුවර්තනය වී ඇතත්, උරහිස උඩින් විසි කිරීමේ සංකීර්ණ හා දුෂ්කර කාර්යය සඳහා තවමත් අස්ථිවිද්යාත්මක වෙනස්කම් ගොඩනැගී නොතිබිණි (Larson et al. 2007). එපමණක් නොව, හෝමෝ ඉරෙක්ටස් මානවයා හා සම්බන්ධ විශේෂිත ප්රක්ෂේපණ ආයුධ (හෙල්ල වැනි) පුරාවිද්යා සන්දර්භය තුළ දක්නට නොමැත.
විශේෂයෙන් ඉරෙක්ටස් හට උරුම වූ දිගු පාද, දිගු දුරක් ධාවනය කිරීමේ හැකියාව තිළිණ කර තිබූ අතර, ඒ මගින් දඩයම් කාර්යයයේ දී විශාල වාසියක් වන්නට ඇති බවට සැකයක් නැත. තවදුරටත්, ෆොසිල වාර්තාවේ එන දිගු ගාත්රා, පටු නම්යශීලී ඉණ සහ කෙටි නළල වැනි ලාක්ෂණික සලකුණු එවැනි දැඩි විඳදරාගැනීමේ හැකියාවක් අවශ්ය වූ ක්රියාකාරකම් සඳහා අනුවර්තනය වී ඇති බව පෙනේ (Bramble & Lieberman 2004; Lieberman et al. 2009). Turkana විලට නැගෙනහිරින් පිහිටි Ileret ප්රදේශයෙන් සංරක්ෂණය වී ඇති ෆොසිල පා සටහන් පෙන්නුම් කරන්නේ ඉරෙක්ටස් ගේ පාද ශරීරයේ අනෙකුත් කොටස් මෙන්ම ඇවිදීමට සහ ධාවනය සඳහා හොඳින් අනුවර්තනය වී තිබුණු බව ය (Lieberman et al. 2009; Bennett et al. 2009). මෙම සියලු ආහාර සෙවීමේ ක්රියාකාරකම් විවෘත, හිරු රශ්මියෙන් දැවෙන සැවානා මත සිදු වීමත් සමඟ, ඉරෙක්ටස් හට පරිසරයේ උෂ්ණත්වය නියාමනය කිරීම අත්යවශ්ය කාරණයක් විය. සිරුරේ ලොම් ගැලවී යාමෙන් දැඩි දහඩිය සිසිලනය වීම ආරම්භ වන්නට ඇති බව විද්වත් මතයයි (Jablonski 2004).

සමාලෝචනය..
අප්රිකාවේ හෝමෝ ගණයේ මූලාරම්භය, පාර්ශවික ද්විපාදකරණයට (partial bipedalism) නතු වූ වානර මානවයින්ගේ සිට ප්රාථමික, විශාල මොළයක් ඇති, ශිලා මෙවලම් සාදන, බොහෝ දුර ගමන් කළ හැකි, මාංශ අනුභව කරන්නන් වෙත අවධානය යොමු කරවයි. මානව කුලයේ මෙම මුල් කොටස විශේෂ තුනකින් නියෝජනය වේ: එනම්, හෝමෝ හැබිලිස්, හෝමෝ රුඩොල්ෆෙන්සිස් සහ හෝමෝ ඉරෙක්ටස් ය. හෝමෝ හැබිලිස් මානවයා විසින් තවදුරටත් ඔස්ට්රොලෝපිතිකස් ජාන සමග සම්බන්ධ කරන ප්රාථමික ලක්ෂණ රඳවා ගත් නමුත්, ඔහු පළමු ශිලාමෙවලම් නිර්මාතෘ ලෙස අවිවාදයෙන් පිළිගැනේ. හැබිලිස් මානවයා හා සසඳන විට සාපේක්ෂ වශයෙන් විශාල මොළයක් සහ විශාල දත් ඇති බව හැර හෝමෝ රුඩොල්ෆෙන්සිස් මානවයා ගැන දන්නේ අල්ප වශයෙන් වන අතර, එම විශේෂය හා මුල් පුර්වජයින් ජීවත් වූ කාලය හා අවකාශය අතිච්ඡාදනය වී ඇත. විශාල මොළයක් ඇති හෝමෝ ඉරෙක්ටස් මානවයාගේ පුරාණජීවවිද්යාව සහ පරිණාමය පිළිබඳ අපගේ අවබෝධය, ඒ සම්බන්ධයෙන් පොහොසත් ෆොසිල වාර්තාව මත රදා පවතී. හෝමෝ ඉරෙක්ටස් මානවයා යනු අනිවාර්යෙන් ම ප්රථම වරට ඍජු කායවිලාසයක් හා සම්පූර්ණයෙන්ම ද්විපාදකරණයට ලක් වූ ශරීරයක් සහිත වන්නා වන අතරම, අප්රිකාවෙන් පිටත විසිරී ගිය මානව පරම්පරාවේ පළමුවැනි සාමාජිකයා ද වේ.
මනුෂ්යත්වයේ කතාන්දරය: මානව පරිණාමය
- මානව පරිණාමයේ ගමන 1: මනුෂ්යත්වයේ ආරම්භය
- මනුෂ්යත්වයේ මූල බීජ: මුල් වානර මානවයින් හා ඔස්ට්රලෝපිතිකස් ගණය
- මුල් හෝමෝ සාමාජිකයින්: හෝමෝ හැබිලිස් හා හෝමෝ ඉරෙක්ටස්
- හෝමෝ නියැන්ඩතාල් මානවයා
- මානව නූතනත්වය: හෝමෝ සේපියන් මානවයාගේ පහළ වීම
- මානව විවිධත්වය හා නව මානව විශේෂ
- මානවයා අප්රිකාවෙන් පිටවී යාම
- මානවයා නවලොව ජනාවාසකරණය කිරීම (අමරිකාව හා ඕස්ට්රේලියාව)
- නියෝලිතික විප්ලවය හා මුල් කෘෂිකර්මයේ නැගීම
කෙටි වාර්තාමය චිත්රපට
- Homo Habilis – Ancient Human
- Homo Erectus: The First Homo Sapiens?
- Primeval Human – Homo Habilis – Early Toolmakers
- The First Humans | Homo Habilis
- Homo Erectus – Ancient Human
- A Brief History Of The Most Successful Human Species: Homo Erectus
References
Arambourg, C. 1955. A recent discovery in human paleontology: Atlanthropus of ternifine (Algeria). American Journal of Physical Anthropology 13: 191–201.
Arambourg, C. & P. Biberson. 1956. The fossil human remains from the Paleolithic site of Sidi Abderrahman (Morocco). American Journal of Physical Anthropology 14: 467–89.
Baur, G. 1895. Pithecanthropus erectus eine menschenähnliche Übergangsform aus Java von Eug. Dubois, Militärarzt der niederländisch-indischen Armee mit zwei Tafeln und drei in den Text gedruckten Figuren. The Journal of Geology 3: 237–38.
Bennett, M.R. et al. 2009. Early Hominin Foot Morphology Based on 1.5-Million-Year-Old Footprints from Ileret, Kenya. Science 323: 1197–1201.
Black, D. 1927. Further Hominid Remains of Lower Quaternary Age from the Chou Kou Tien Deposit. Nature 120: 954–954.
Black, D. 1934. III. The croonian lecture. – On the discovery, morphology, and environment of Sinanthropus pekinensis. Philosophical Transactions of the Royal Society of London. Series B, Containing Papers of a Biological Character 223: 57–120.
Bobe, R. 2004. The expansion of grassland ecosystems in Africa in relation to mammalian evolution and the origin of the genus Homo. Palaeogeography, Palaeoclimatology, Palaeoecology 207: 399–420.
Bobe, R. & M.G. Leakey. 2009. Ecology of Plio-Pleistocene Mammals in the Omo—Turkana Basin and the Emergence of Homo, in : 173–84.
Bramble, D.M. & D.E. Lieberman. 2004. Endurance running and the evolution of Homo. Nature 432: 345–52.
Braun, D.R., T. Plummer, P. Ditchfield, J. v. Ferraro, D. Maina, L.C. Bishop & R. Potts. 2008a. Oldowan behavior and raw material transport: perspectives from the Kanjera Formation. Journal of Archaeological Science 35: 2329–45.
Braun, D.R., J.C. Tactikos, J. v. Ferraro, S.L. Arnow & J.W.K. Harris. 2008b. Oldowan reduction sequences: methodological considerations. Journal of Archaeological Science 35: 2153–63.
Braun, D.R., J.W.K. Harris, N.E. Levin, J.T. McCoy, A.I.R. Herries, M.K. Bamford, L.C. Bishop, B.G. Richmond & M. Kibunjia. 2010. Early hominin diet included diverse terrestrial and aquatic animals 1.95 Ma in East Turkana, Kenya. Proceedings of the National Academy of Sciences 107: 10002–7.
Brown, F., J. Harris, R. Leakey & A. Walker. 1985. Early Homo erectus skeleton from west Lake Turkana, Kenya. Nature 316: 788–92.
Capasso, L., E. Michetti & R. D’Anastasio. 2008. A Homo erectus hyoid bone: possible implications for the origin of the human capability for speech. Collegium antropologicum 32: 1007–11.
Clarke, R.J., F.C. Howell & C.K. Brain. 1970. More evidence of an advanced hominid at Swartkrans. Nature 225: 1219–22.
Day, M.H. 1971. Postcranial remains of Homo erectus from Bed IV, Olduvai Gorge, Tanzania. Nature 232: 383–87. https://doi.org/10.1038/232383a0.
Day, M.H. & R.E. Leakey. 1973. New evidence of the genus Homo from East Rudolf, Kenya. I. American journal of physical anthropology 39: 341–54.
Day, M.H., R.E. Leakey, A.C. Walker & B.A. Wood. 1976. New hominids from East Turkana, Kenya. American journal of physical anthropology 45: 369–435.
deMenocal, P.B. 2004. African climate change and faunal evolution during the Pliocene–Pleistocene. Earth and Planetary Science Letters 220: 3–24.
Domínguez-Rodrigo, M., T.R. Pickering, S. Semaw & M.J. Rogers. 2005. Cutmarked bones from Pliocene archaeological sites at Gona, Afar, Ethiopia: implications for the function of the world’s oldest stone tools. Journal of human evolution 48: 109–21.
Dunsworth, H.M. 2010. Origin of the Genus Homo. Evolution: Education and Outreach 3: 353–66.
Gabunia, L. & A. Vekua. 1995. A Plio-Pleistocene hominid from Dmanisi, East Georgia, Caucasus. Nature 373: 509–12.
Gabunia, L. et al. 2000. Earliest Pleistocene hominid cranial remains from Dmanisi, Republic of Georgia: taxonomy, geological setting, and age. Science (New York, N.Y.) 288: 1019–25.
Haeusler, M. & H.M. McHenry. 2004. Body proportions of Homo habilis reviewed. Journal of Human Evolution 46: 433–65.
Heinzelin, J. de, J.D. Clark, T. White, W. Hart, P. Renne, G. WoldeGabriel, Y. Beyene & E. Vrba. 1999. Environment and Behavior of 2.5-Million-Year-Old Bouri Hominids. Science 284: 625–29.
Holloway, R.L. 1983. Human paleontological evidence relevant to language behavior. Human neurobiology 2: 105–14.
Hughes, A.R. & P. v Tobias. 1977. A fossil skull probably of the genus Homo from Sterkfontein, Transvaal. Nature 265: 310–12.
Jablonski, N.G. 2004. The Evolution of Human Skin and Skin Color. Annual Review of Anthropology 33: 585–623. https://doi.org/10.1146/annurev.anthro.33.070203.143955.
Johanson, D.C. et al. 1987. New partial skeleton of Homo habilis from Olduvai Gorge, Tanzania. Nature 327: 205–9.
Larson, S.G. 2009. Evolution of the Hominin Shoulder: Early Homo, in The first humans—origin and early evolution of the genus Homo: 65–75.
Larson, S.G., W.L. Jungers, M.J. Morwood, T. Sutikna, Jatmiko, E.W. Saptomo, R.A. Due & T. Djubiantono. 2007. Homo floresiensis and the evolution of the hominin shoulder. Journal of human evolution 53: 718–31.
Leakey, L.S.B. 1961. New Finds at Olduvai Gorge. Nature 189: 649–50.
Leakey, L.S.B., P. v. Tobias & J.R. Napier. 1964. A New Species of The Genus Homo From Olduvai Gorge. Nature 202: 7–9.
Leakey, M.D. 1971. Discovery of postcranial remains of Homo erectus and associated artefacts in Bed IV at Olduvai Gorge, Tanzania. Nature 232: 380–83.
Leakey, R.E. 1973a. Further evidence of lower pleistocene hominids from East Rudolf, North Kenya, 1972. Nature 242: 170–73.
—. 1973b. Evidence for an advanced plio-pleistocene hominid from East Rudolf, Kenya. Nature 242: 447–50.
Lieberman, D.E., D.M. Bramble, D.A. Raichlen & J.J. Shea. 2009. Brains, Brawn, and the Evolution of Human Endurance Running Capabilities, in : 77–92.
Lordkipanidze, D. et al. 2005. Anthropology: the earliest toothless hominin skull. Nature 434: 717–18.
—. 2007. Postcranial evidence from early Homo from Dmanisi, Georgia. Nature 449: 305–10.
Maslin, M.A. & M.H. Trauth. 2009. Plio-Pleistocene East African Pulsed Climate Variability and Its Influence on Early Human Evolution, in : 151–58.
Miller, J.A. 1991. Does brain size variability provide evidence of multiple species in Homo habilis? American journal of physical anthropology 84: 385–98.
Movius, H.L. 1948. The Lower Palaeolithic Cultures of Southern and Eastern Asia. Transactions of the American Philosophical Society 38: 329.
Potts, R. 1998a. Environmental hypotheses of hominin evolution. American journal of physical anthropology Suppl 27: 93–136.
Potts, R. 1998b. Variability selection in hominid evolution. Evolutionary Anthropology: Issues, News, and Reviews 7: 81–96.
Reed, K.E. 1997. Early hominid evolution and ecological change through the African Plio-Pleistocene. Journal of Human Evolution 32: 289–322.
Reed, K.E. & S.M. Russak. 2009. Tracking Ecological Change in Relation to the Emergence of Homo Near the Plio-Pleistocene Boundary, in : 159–71.
Rightmire, G.P., D. Lordkipanidze & A. Vekua. 2006. Anatomical descriptions, comparative studies and evolutionary significance of the hominin skulls from Dmanisi, Republic of Georgia. Journal of Human Evolution 50: 115–41.
Roche, H., R.J. Blumenschine & J.J. Shea. 2009. Origins and Adaptations of Early Homo: What Archeology Tells Us, in : 135–47.
Ruff, C.B., E. Trinkaus & T.W. Holliday. 1997. Body mass and encephalization in Pleistocene Homo. Nature 387: 173–76.
Sartono, S. 1972. Discovery of Another Hominid Skull at Sangiran, Central Java. Current Anthropology 13: 124–26.
Semaw, S. et al. 2003. 2.6-Million-year-old stone tools and associated bones from OGS-6 and OGS-7, Gona, Afar, Ethiopia. Journal of human evolution 45: 169–77.
Sonakia, A. & K.A.R. Kennedy. 1985. Skull Cap of an Early Man from the Narmada Valley Alluvium (Pleistocene) of Central India. American Anthropologist 87: 612–16. https://doi.org/10.1525/aa.1985.87.3.02a00060.
Sponheimer, M. & J.A. Lee-Thorp. 2009. Biogeochemical Evidence for the Environments of Early Homo in South Africa, in : 185–94.
Susman, R.L. 2008. Brief communication: Evidence bearing on the status of Homo habilis at Olduvai Gorge. American Journal of Physical Anthropology 137: 356–61.
Tobias, P. v. 2009. Homo habilis—A Premature Discovery: Remembered by One of Its Founding Fathers, 42 Years Later, in : 7–15.
Ungar, P. 2004. Dental topography and diets of Australopithecus afarensis and early Homo. Journal of human evolution 46: 605–22.
Ungar, P.S. 2012. Dental Evidence for the Reconstruction of Diet in African Early Homo. Current Anthropology 53: S318–29.
Ungar, P.S. & R.S. Scott. 2009. Dental Evidence for Diets of Early Homo, in : 121–34.
Ungar, P.S., F.E. Grine, M.F. Teaford & S. el Zaatari. 2006. Dental microwear and diets of African early Homo. Journal of Human Evolution 50: 78–95.
Weidenreich, F. 1944. Giant Early Man from Java and South China. Science 99: 479–82.
Wood, B. 1992. Origin and evolution of the genus Homo. Nature 355: 783–90.
Wood, B. & M. Collard. 1999. The changing face of genusHomo. Evolutionary Anthropology: Issues, News, and Reviews 8: 195–207.